

1 Department of Foreign Languages and Linguistics, Shiraz University, Shiraz, 7194685115, Iran
2 DANA Brain Health Institute, Iranian Neuroscience Society, Fars Chapter, Shiraz, 7183611557, Iran
3 Neuroscience Laboratory-NSL (Brain, Cognition and Behavior), Department of Neuroscience, School of Advanced Medical Sciences and Technologies, Shiraz University of Medical Sciences, Shiraz, 7184737798, Iran
4 Department of Neuroscience, School of Advanced Medical Sciences and Technologies, Shiraz University of Medical Sciences, Shiraz, 7184737798, Iran
5 Academy of Health, Senses Cultural Foundation, Sacramento, CA, 95826, USA
Abstract
To evaluate transcranial direct current stimulation-induced changes in resting state quantitative EEG and cognitive-verbal performance of second language learners, 16 healthy individuals were randomly recruited to sham and real transcranial direct current stimulation groups receiving eight sessions of second language instruction accompanied by a 2 mA transcranial direct current stimulation over Broca's area with the cathode placed over the left arm. Quantitative EEG was recorded during the resting state after the stimulation session and second language instruction. Reduced theta activity at Fp1, F7, F3, and T5 caused by the stimulus current was reported. Multisession stimulation resulted in a significant increase in current density for beta power (25 Hz) in the language network. Cognitive-verbal pre-post stimulation performances suggest that anodal vs. sham transcranial direct current stimulation significantly improved the subjects test score on digit span, a cognitive-verbal ability. It is concluded that transcranial direct current stimulation of Broca's area increase cognitive-verbal performance by modulating brain electrical activity in language-related regions.
Keywords
- qEEG
- transcranial direct current stimulation
- Broca's area
- cognitive-verbal function
- current density
A comprehensive body of knowledge has reported the potency of brain stimulation for non-therapeutic purposes including cognitive performance enhancement such as improving threat detection ability (Bullard et al., 2011), enhancing verbal memory for learning nouns (Elmer et al., 2009), upgrading the recognition memory in patients suffering from Alzheimer (Ferrucci et al., 2008), increasing lexical retrieval and naming and decreasing response time (Fertonani et al., 2010), facilitation of implicit learning (Kincses et al., 2004), improving declarative memory (Marshall et al., 2004), boosting working memory cognitive training and skill acquisition (Martin et al., 2014), and promoting working memory (Ohn et al., 2008). Anodal and cathodal transcranial direct current stimulation (tDCS) result respectively in depolarization and hyperpolarization (Filmer et al., 2014) through changing cortical excitability caused by application of a weak direct current. Modulation of regional electrical activity (Keeser et al., 2011), local blood volume (Baudewig et al., 2001) and cortical neurotransmitters (Stagg et al., 2009) through tDCS have also been reported. Excitation induced by tDCS has been shown to decrease delta (1-4 Hz) and theta (4-8 Hz) EEG power (Keeser et al., 2011) while increasing beta (12-25 Hz) and gamma (30-40 Hz) activity (Hlinka et al., 2010; Keeser et al., 2011). A study by Keeser and his colleagues (Keeser et al., 2011) attempted to evaluate changes in electrical activity following anodal tDCS over the left dorsolateral prefrontal cortex (DLPFC) and cathodal tDCS over the right supraorbital region along with any subsequent impact on working memory function. Results of the above investigation showed increased and decreased neuronal electrical activity of beta and delta-theta frequencies, respectively. They reflect the vigilance changes that resulted in more favorable performance in subsequent working memory tasks.
Functional magnetic resonance imaging (fMRI) and positron emission tomography (PET) results have shown that anodal tDCS increases both blood-oxygen-level dependent (BOLD) (Baudewig et al., 2001) and cortical hemodynamic activity (Lang et al., 2005). Moreover, magnetic resonance spectroscopy results have shown changes in GABAergic inhibition due to anodal brain stimulation (Stagg et al., 2009). Thus, it has been proposed that tDCS-induced changes in electrical and metabolic activities as well as neurotransmitters may enhance cognitive functions and learning (Clark et al., 2012). Further, Broca's area has been shown to have a crucial role for language processing (Bernal et al., 2015; Cappelletti et al., 2008; Hagoort, 2005; Kunert et al., 2015; Novick et al., 2005, 2010; Smits et al., 2006) and to be involved in higher order complex cognitive functions (Novick et al., 2010). According to Hagoort, 2005, the area is not language-specific and even within language-related functions, it plays more than one role as it can be activated during field independent tasks (i.e. tasks including details separated from the surrounding context), recognition functions and movement.
The left lateral frontopolar area 1 (Fp1), dorsolateral prefrontal cortex (F3), left inferior frontal lobule (F7), and the left temporal cortex (T5) are language related regions of interest (ROI). Electrical activity in areas F3 and F7 originates from frontal brain regions involved in language processing (Weiss and Mueller, 2003). Additionally, occipital and fronto-polar (Fp) areas are known to be active during word processing (Weiss et al., 2000) and Wernicke's area (T5) is also known to be involved in language comprehension and production (Binder, 2015). However, previous studies have not investigated the possible effect of tDCS in Broca's area on electrophysiological changes within the language network (i.e. key areas including FP1, F3, F7 and T5). Therefore, this study combined tDCS and quantitative electroencephalography (qEEG) to further elucidate the neurophysiological mechanisms underlying tDCS and any associated stimulation induced cognitive-verbal changes. It was hypothesized that neurophysiological changes following brain stimulation-induced neuromodulation could positively affect the cognitive-verbal performance of second language (L2) students. Moreover, the subsequent alteration in resting state qEEG dynamics was also hypothesized to be associated with the improvements predicted for cognitive-verbal performance.
Sixteen (Kasahara et al., 2013) right-handed (Oldfield, 1971) healthy subjects (based on an interview) aged 18 to 31 years were included in the study. Their native language was Persian and they had basic proficiency in English. Through a design approved by the Ethics Committee of Shiraz University of Medical Sciences (code 97901-1089), subjects were randomly assigned to two parallel groups receiving either a 2 mA sham or real tDCS, respectively, following eight sessions of English instruction as a second language (L2). Table 1 gives the demographic information of the subjects.
| Real tDCS arm (n = 8) | Sham tDCS arm (n = 8) | |
|---|---|---|
| Male/female | 3/5 | 3/5 |
| Mean age in years | 23.8 ± 4.5 | 23.5 ± 3.6 |
| Mean years of formal education | 15.50 ± 2 | 14.25 ± 1.5 |
The study was designed as a double-blind, sham-real tDCS experiment. This study had no control group (i.e. the group of subject receiving neither sham nor real tDCS intervention). The study groups comprised real and sham tDCS groups which underwent identical language training. A non-researcher assigned subjects to either group, with subjects unaware of their group assignment. Data were collected over ten daily sessions each separated by 24 hours. Consent forms were collected and Cambridge Brain Sciences-Cognitive Platform (CBS-CP) trials were done during the first session prior to either sham or real tDCS (pre-intervention). In sessions two to nine, both groups underwent their respective 2 mA real or sham tDCS (tDCS intervention) for 20 minutes while receiving L2 grammar instruction on eight simple to complex grammatical structures chosen based on Common European Framework of Reference for Languages (CEFR) levels. After sessions 2 and 9, a five minute eyes-open resting state qEEG was recorded. In the last session, subject performance on CBSCP verbal trials, which functioned as the post-intervention performance, was obtained. To ensure that CBS-CP performance reflected cognitive ability, subjects received CBS-CP training upon sessions 1 and 10 before and after receiving either sham or real tDCS (pre- and post-tDCS intervention).
A tDCS device (NeuroStim2) was used to deliver a 2 mA electrical current for 20 minutes (Holland et al., 2011) over eight sessions. The current was bounded by a 30 second stimulus ramp up and down. In several previous studies a two mA current was initially tested (Fertonani et al., 2010; Fiori et al., 2014; Holland et al., 2011; Kamali et al., 2019a). Although the same current magnitude was employed in this investigation, further studies could address whether a lower current might achieve similar results. Due to limitation in sample size, number of test groups and the longitudinal nature of this study, the 2mA current was the only stimulus applied. Each 9 cm2 electrode was positioned over a 3 × 3 cm salinesoaked (NaCl 150 mM) sponge. The anode and cathode electrodes were placed over Broca's area (44/45 Brodmann area) and on the left arm (over the deltoid muscle), respectively. Electrical stimulation, was applied for 20 minutes to subjects in the real tDCS group, whereas for the sham group, stimulation was switched off after 30 seconds, a period of time sufficient to initiate a sensation of stimulation.
The effect of stimulation on resting state qEEG was recorded (EEG 3840, Mitsar) using a 19-electrode EEG cap, with electrodes placed according to the International 10/20 system (reference channel Cz). Skin impedance was < 10 kΩ and the sampling rate was 150 Hz. Before analysis, eye and muscle artifacts detected visually were excluded. Following signal preprocessing (Mitsar amplifier 10/70-201, Russia) by built-in amplifier software (WinEEG, version 2.127.98) data were further visually analyzed by two EEG experts to exclude distorted and noisy signal segments. Signal components that exhibited amplitude overshot and EEG distortion due to muscle, blink and movement artifacts were also removed. This was visually crossed-checked by two electrophysiologists. Six-second data epochs were analyzed using NeuroGuide software (v. 3.0.2 2001-2018 Applied Neuroscience Inc. USA). EEG absolute power (µV2) of the theta (4-8 Hz), alpha (8-12 Hz), beta (12-25 Hz) and high beta ( 25-30 Hz) frequency band were compared for language ROI (Fp1, F3, F7, and T5) in the sham and real tDCS groups for sessions one and eight. The NeuroGuide frequency spectrum suggested that the language network (Brodmann areas 22, 39, 40, 41, 42, 44, 45) current spectral density (CSD) was predominantly a 25 Hz oscillation in the resting state qEEG. Consequently, the mean CSD z-score value of the seven language-related Brodmann areas was calculated and compared for the real and sham tDCS groups.
To assess the cognitive-verbal ability of the participants, CBS- CP was applied. This is one of the most widely studied and validated platforms for profiling cognitive aptitude. Its capacity for comparative analysis and z-score reporting based on normative data derived from a database comprising over 40,000 subjects is a key advantage of the test (Hampshire et al., 2012). As a cultureblind media-rich digital platform, CBS-CPr is widely used in clinical and applied neurocognitive research. Alternative software includes CANTAB (Sweeney et al., 2000), IVA (Wesnes et al., 2013) and CogniPlus (Hagovska et al., 2016). CBS-CP was chosen on the basis of its robust validity and ease of use as well as its wide use in previously published reports (Ferreira et al., 2015; Kamali et al., 2019b; Loughman et al., 2014; Owen et al., 2010).
The verbal ability trials of CBS-CP that were employed here included Digit Span, Double Trouble and Grammatical Reasoning (see Hampshire et al., 2012 Digit Span (Bor et al., 2004)) primarily relies on verbal ability and to lesser extent short-term working and digit memory. Since memorizing items in a specific word order is required for this task, it can be useful for quantifying understanding of long and complex sentences. The task can be associated with storage and rehearsal of words in what is referred to as the phonological loop. In the test a number of digits appear sequentially on a screen and subjects are required to memorize them in the given order. They are then requireded to record the digits in the same order. Double Trouble (Stroop, 1935) is a verbal response inhibition task that aims to increase concentration in the presence of distracting information. This task originates from the Stroop effect where the color of a word representing another color must be named. Because of the primary word recognition mechanism, this is difficult. Three words are shown on a screen: for example, two words at the bottom are "RED" and "BLUE" and one at the top changes randomly. Subjects are required to choose either "RED" "BLUE" based on the color of the random word that appears at the top of the screen. The task can be effective for activities where a person must ignore distracting information to focus on important information. Grammatical Reasoning (Hampshire et al., 2012) is a verbal reasoning task that evaluates valid understanding of words. The task is challenging due to complex passive and negative sentences. Two geometrical shapes as well as a sentence appear on the screen. The participants are required to select "True" or "False" according to the correctness of the sentence in describing the relationship between the shapes. Accuracy and speed matter for obtaining high scores. The task is mainly used for evaluating short- and long-term cognitive impairment and is effective for accurately quantifying understanding of language.
To compare the effects of L2 grammar instruction accompanied by either sham or real tDCS, the subjects in each of the two groups were required to perform the trials once before and once after the tDCS intervention.
To test whether there is any statistically significant difference between the resting state absolute power of language ROI in different frequency bands obtained for the sham a tDCS groups, a one-way repeated measure multivariate analysis of variance (MANOVA) was conducted. This test was used as a one-way analysis of variance (ANOVA) evaluates only one dependent variable at a time, whereas, the MANOVA is designed to test differences in multiple dependent variables over time and between different treatments. Here, the within-subject variable was absolute power and the between-subject variables were region, tDCS type, and frequency band.
To explore the differences between the language network CSDs of the z-score between the two groups over the course of treatment, a mixed between-within subjects ANOVA was run. Compared to an independent-sample t-test, this type of ANOVA compares the mean difference between groups separated by two independent variables, one within-subjects and one between-subjects. For this study, the between-subject variable was the type of brain stimulation (sham or real tDCS) and the within-subjects variable was time (pre- and post-tDCS intervention). A series of one-way between-group MANOVAs were performed to investigate the difference between the sham and real tDCS groups in terms of CBSCP verbal trials. Compared to the one-way ANOVA measuring one dependent variable, a one-way MANOVA evaluates the difference between groups on the basis of more than one dependent variable. In this study, there were three dependent variables, the results of the Digit Span, Double Trouble, and Grammatical Reasoning tests. To avoid Type 1 error due to multiple comparisons in one-way MANOVAs, a Bonferroni adjustment was made for the results of tests of the between-subject effects (Pallant, 2013). The SPSS statistical package (Version 23.0.0, Copyright©IBM, 2015) was used for data analysis and a P-value < 0.05 was considered significant.
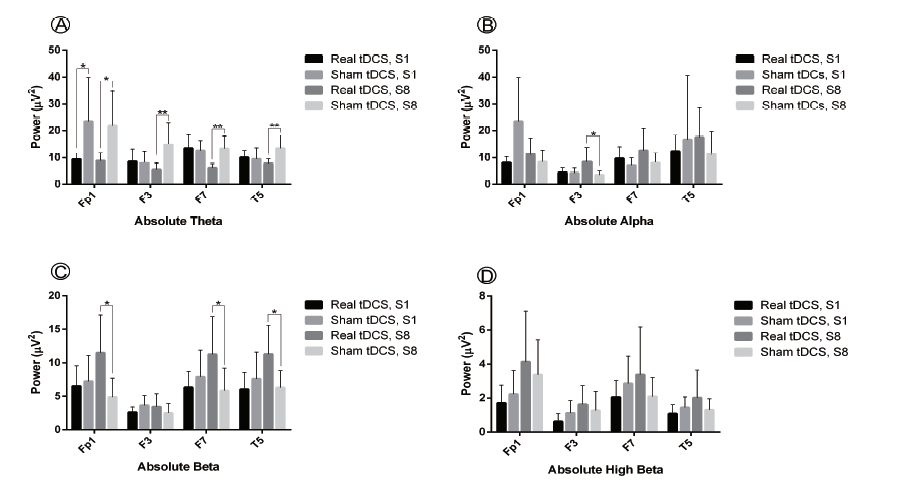
A one way repeated-measures MANOVA was used to explore how sham and tDCS group qEEG resting state frequency absolute power (µV2) in four language-related ROI were affected by tDCS over time. Within-subject variables were the absolute power values of sessions one and eight and the between-subject variable was region (Fp1, F3, F7 and T5). Intervention type (sham or real tDCS) and frequency band (theta, alpha, beta, high beta) were covariates. This analysis showed significant interaction between time and tDCS type (Wilk's Λ = 0.983, F(1, 250) = 4.36, P = 0.038, ηp2 = 0.017), suggesting a significant difference between real and sham tDCS over the course of treatment. Similarly, as for the time, the effect was statistically significant (Wilk's Λ = 0.977, F(1, 250) = 6.00, P = 0.015, ηp2 = 0.023). However, there was not a significant interaction between time and frequency ( Wilk's Λ = 0.998, F(1, 14) = 524, P = 0.470, ηp2 = 0.002), suggesting that treatment time did not affect frequency band absolute power in the two groups. Likewise, no significant interaction was detected between time and region (Wilk's Λ = 0.977, F(1, 250) = 0.238, P = 0.870, ηp2 = 0.003). As the result revealed, tDCS over Broca's area significantly decreased theta power at the Fp1 electrode (t(14) = -2.40, P = 0.031) in session one. Regarding session eight, tDCS resulted in a significant decrease in theta power at the Fp1, F3, F7 and T5 electrodes (Fp1: t(14) = -2.79, P = 0.014; F3: t(14) = -3.03, P = 0.009; F7: t(14) = -4.24, P = 0.001; T5: t(14) = -3.20, P = 0.006) and increase of F3 alpha as well as Fp1, F7 and T5 beta power (Fp1: t(14) = 2.96, P = 0.010; F3: t(14) = 2.61, P = 0.02; F7: t(14) = 2.34, P = 0.034; T5: t(14) = 2.84, P = 0.013). Fig. 1 gives the qEEG absolute power at four ROI for both sham stimulation and tDCS in theta, alpha, beta and high beta bands in session one and eight.
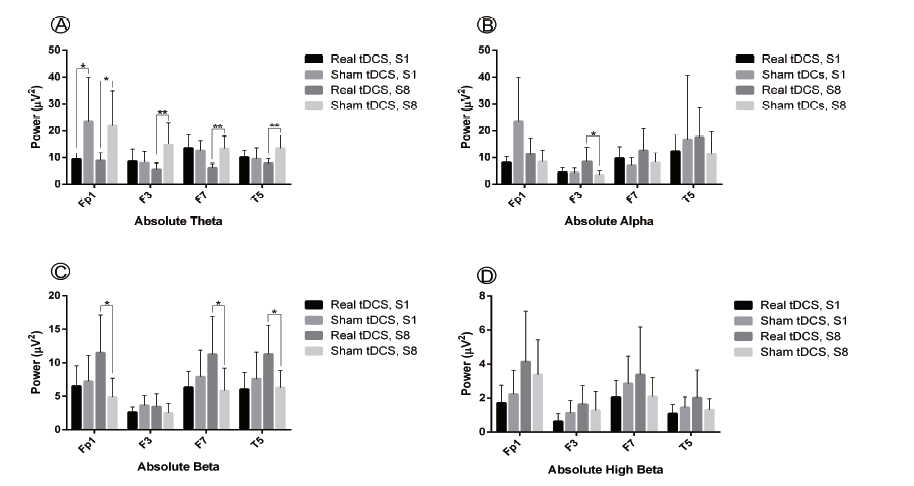 Figure 1.
Figure 1.Effect of real vs. sham tDCS on linguistic ROI absolute power (µV2) for frequency bands theta (4-8 Hz), alpha (8-12 Hz), beta (12--30 Hz) and high beta (25-30 Hz) in sessions 1 (S1) and 8 (S8) of tDCS. Multi-session real vs. sham tDCS induced a significant decrease in theta power in linguistic ROI and significantly increased beta power at F7 and T5. Statistical significance assumed for P < 0.05, results given as mean ± standard error of the mean (SEM).
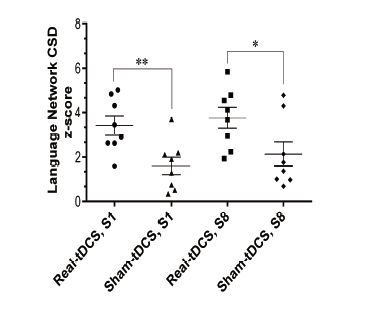
To evaluate how the language network CSD z-score was affected by tDCS, the average score of language network (Brodmann areas 22, 39, 40, 41, 42, 44, 45) CDS z-score beta power (25 Hz oscillation) in session one and eight were compared in the sham and real tDCS groups through a mixed between-within subjects ANOVA. Homogeneity of both inter-correlations and variances revealed no detectable differences. No significant interaction was found between time and tDCS intervention type (i.e. sham and real) (Wilk's Λ = 0.997, F(1, 14) = 0.046, P = 0.833, ηp2 = 0.003), suggesting that the treatment time did not affect the type of tDCS intervention (sham or real). Similarly, as for time, the effect was not statistically significant (Wilk's Λ = 0.933, F (1, 14) = 1.01, P = 0.331, ηp2 = 0.067). However, there was a significant main effect for tDCS intervention type (i.e. sham or real) (F(1, 14) = 12.1, P = 0.004, ηp2 = 0.465), suggesting a significant difference in the effectiveness of real and sham tDCS in changing the language network CSD z-score. Sham stimulation vs. tDCS significantly increased the language network CSD z-score upon rest both in session one (t(14) = 3.13, P = 0.007, η2 = 0.410) and session eight (t(14) = 2.25, P = 0.041, η2 = 0.265) (Fig. 2). Effects of sham vs. real tDCS-included L2 grammar instruction on language network CSD z-score is demonstrated in Fig. 3.
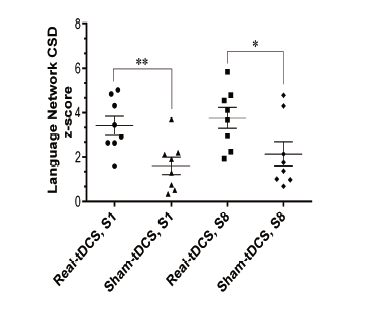 Figure 2.
Figure 2.Dot plots give mean CSD z-score of language network centers at 25 Hz oscillation that showed significant loading during the resting state. Real vs. sham tDCS resulted in a significant mean CSD z-score in sessions 1 (S1) and 8 (S8) during the resting state (P < 0.05). Intra-group comparisons revealed no significant difference between the two group mean values in S1 and S8. Statistical comparison: mean ± standard error of the mean (SEM), significance level: P < 0.05 indicated by asterisk (*).
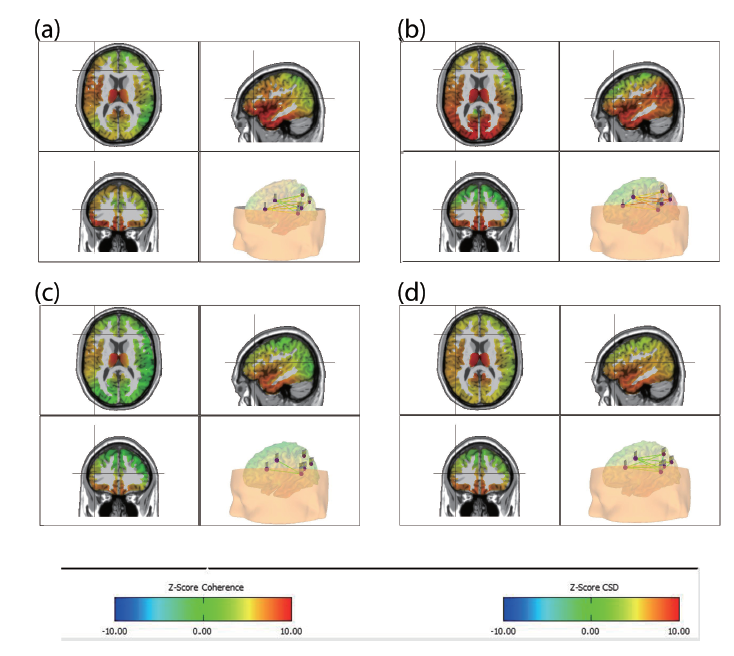
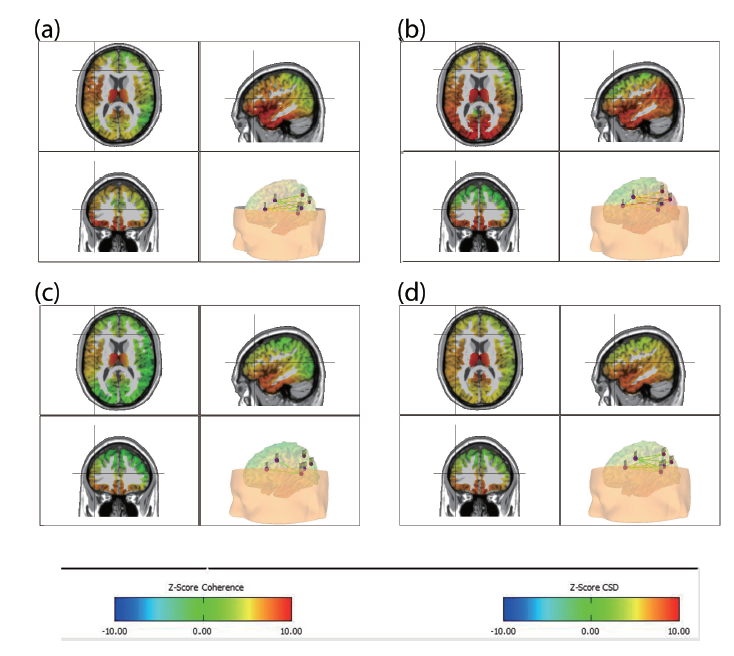 Figure 3.
Figure 3.Average effect of sham vs. real tDCS on the mean language network CSD z-score at 25 Hz for the resting state EEG. Panel a, b: Real tDCS group average CSD z-score after one and eight sessions of tDCS. Panel c, d: Average CSD z-score of the sham group in the first and eight sessions of tDCS.
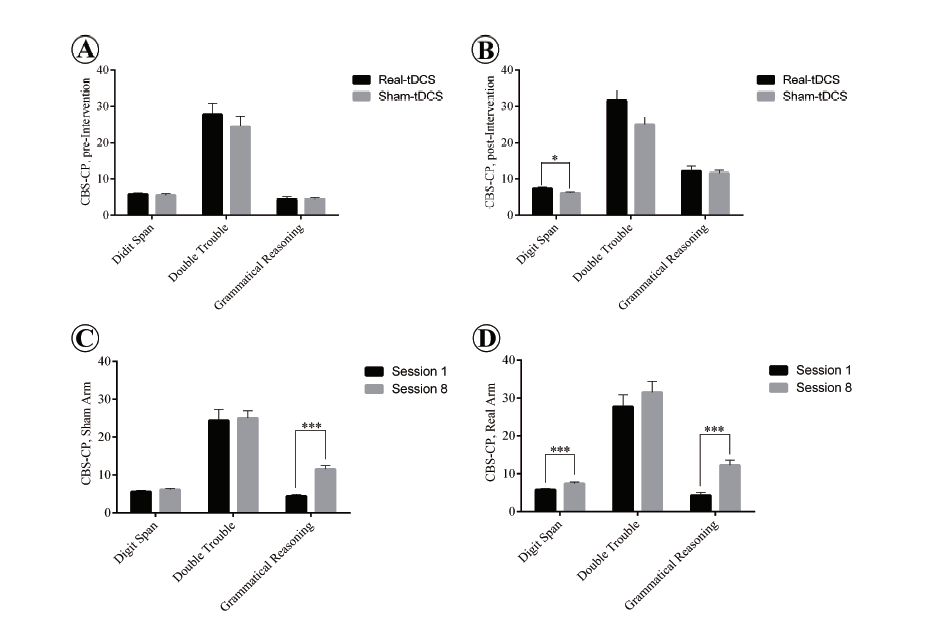
To explore the effect of real and sham tDCS accompanying grammar instruction on the subjects cognitive functions, the two groups performances on verbal trials of the CBS-CP, both before and after the tDCS intervention, were compared through a series of one-way MANOVAs. Subject performance was taken to be the dependent variable in the CBS-CP verbal component (i.e., for the Digit Span, Double Trouble and Grammatical Reasoning tests). The independent variable was the type of tDCS intervention offered, i.e. sham or real tDCS. A check of preliminary assumptions showed no serious departures from normality and linearity or deviations of univariate and multivariate outliers, homogeneity of variance-covariance matrices, and multicollinearity. The results of CBS-CP verbal trials are given in Fig. 4. Results obtained for the pre-intervention scores of the sham and real tDCS groups showed no statically significant difference between them for the combined dependent variables (F(3, 12) = 0.227, P = 0.876; Wilks Λ = 0.946).
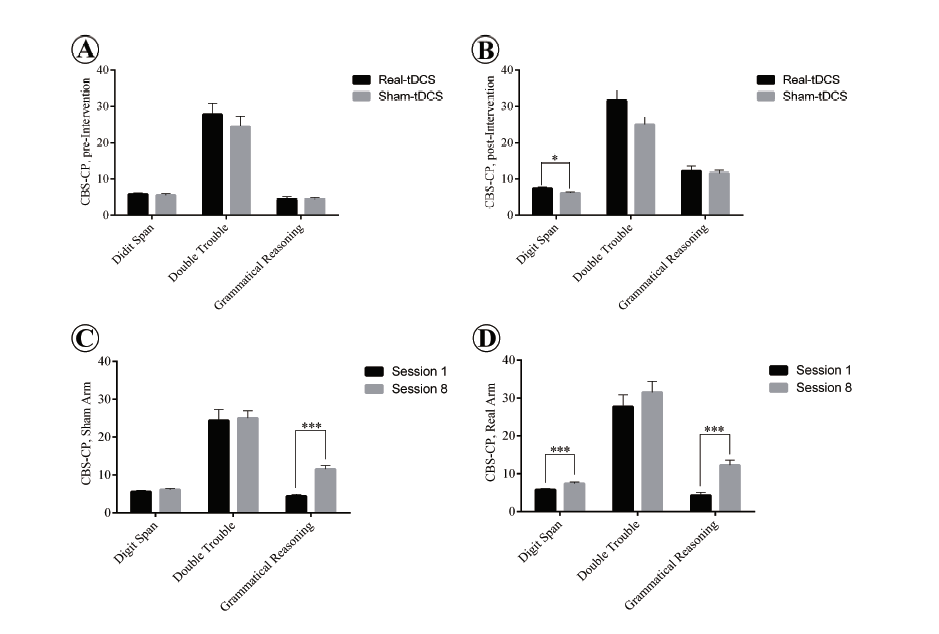 Figure 4.
Figure 4.Subject performance in verbal trials of CBS-CP before and after tDCS intervention. Panel A: No significant difference between the performance of the subjects on three verbal tasks in the sham and real tDCS groups. Panel B: tDCS accompanying L2 grammar instruction significantly improves subject performance in the Digit span task compared with sham stimulation (P < 0.05). Panel C: Significant difference in Grammatical reasoning task from preto post-intervention in the sham group (P < 0.05). Panel D: Significant improvement of the tDCS group from preto post-intervention on Digit Span and Grammatical Reasoning tasks (P < 0.05). Significant P-value 0 .05. Asterisk (*) indicates significant differences, data summarized as mean ± standard error of the mean (SEM).
A statistically significant difference in post-intervention scores was found between the sham and real tDCS groups for the combined dependent variables (F(3, 12) = 4.60, P = 0.023; Wilks Λ = 0.462), and the effect size was found to be large (d = 0.535). Since a significant multivariate main effect for the verbal component was indicated, a test of between-subjects effect was run for each dependent variable. The only statistically significant difference, using a Bonferroni adjusted α = 0.017 (α = 0.5 divided by three to reach an acceptable confidence level for each of the three trials) was found for Digit Span (F(1, 14) = 5.93, P = 0.029, ηp2 = 0.298).
With regard to intra-comparison of group pre- and postintervention performances for the CBS-CP verbal component, a series of one-way MANOVAs was undertaken with verbal trials as the dependent variable and the time (pre- and post-intervention) as the independent variable. Results for the tDCS group showed a statistically significant difference between pre- and post-intervention for the combined dependent variables (F(3, 12) = 12.2, P = 0.001; Wilks Λ = 0.246), Digit Span (F(1, 14) = 8.51, P =0.011, ηp2 = 0.378) and Grammatical Reasoning (F( 1, 14) = 25.1, P < 0.001, ηp2 = 0.642) at a Bonferroni adjusted α = 0.017. Similarly for the sham group, there was a statistically significant difference between pre- and post-intervention on the combined dependent variables (F(3, 12) = 17.8, P < 0.001; Wilks Λ = 0.184). Use of a Bonferroni adjusted α = 0.017 significantly increased the Grammatical Reasoning score from pre- to post-intervention (F(1, 14) = 41.8, P < 0.001, ηp2 = 0.749).
By application of combined behavioral-tDCS intervention, this study attempted to explore electrocortical modulation and cognitive-verbal changes induced by multi-session stimulation of Broca's area. In the case of cognitive performance, a significant benefit of tDCS was reported for Digit Span; however, both sham and real tDCS resulted in a significant improvement in performance upon post-intervention evaluation in Grammatical Reasoning task. The significantly improved performance seen for Digit Span as a cognitive-verbal ability (prompted by sham vs. tDCS) can be justified by fMRI results verifying the predominant role of the left inferior frontal gyrus (IFG) in processing this cognitiveverbal task (Hampshire et al., 2012). Additionally, the evidence could support a role for Broca's area in short-term, digit (Baddeley and Hitch, 1974; Baddeley, 2003), and verbal working memories (Fiebach et al., 2001; Rogalsky et al., 2008).
Results reported here are supported by an earlier study that showed the effectiveness of tDCS over frontal areas in enhancement of verbal memory tasks (Arnold and Nozari, 2017; Ohn et al., 2008) the effectiveness of stimulation of Broca's area for picture naming (Holland et al., 2011), accurate/fast production of tongue twisters (Fiori et al., 2014), and modulating cognitive control (Lupyan et al., 2012). The significant improvement of the two groups in the Grammatical Reasoning task is also supported by an earlier study that showed the effectiveness of anodal tDCS over the left IFG for matching images with descriptions (Lupyan et al., 2012) as the Grammatical Reasoning task also requires subjects to make decisions concerning the fitness of the descriptions for the image stimuli. Results can also be attributed to the L2 grammar instruction. The absence of significant results in the Double trouble test can be attributed to the primary focus of the task being response inhibition and concentration on relevant information rather than language, though being categorized in the verbal component of CBS-CP. Future studies could explore brain stimulation protocols for response inhibition relevant-irrelevant information cognitive tasks.
Another possible explanation for enhanced performance in cognitive-verbal trials could be shortening of reaction time (i.e. improved cognitive agility) following tDCS (Keeser et al., 2011; Nitsche et al., 2003). These results are also supported by a tDCSfMRI study (Holland et al., 2011) that showed the facilitatory effect of Broca area stimulation in reduction of reaction time and improved image naming, presumably due to a reduced BOLD signal. Nonetheless, one aspect which might be criticized is the timedelay for cognitive tasks as they were performed once before and once after the course of treatment on a separate day, not immediately after tDCS. This time delay for the cognitive task might affect subject performance in verbal trials. Further, it has already been shown that repeated sessions of tDCS may result in enduring effects and long-term plasticity (Chesters et al., 2018; Cohen et al., 2010).
TDCS-induced decrease in theta power in Fp1, F3, F7 and T5 confirms former studies (Keeser et al., 2011; Marshall et al., 2004) that reported theta power decreases in frontal, central and parietal regions due to frontal brain stimulation. Results also support another study (Ardolino et al., 2005) which indicated that anodal tDCS and magnetic stimulation reduced theta activity in the stimulated region and connected areas. Moreover, significant increase in beta power in Fp1, F7 and T5 regions confirmed results which showed that cortical excitability through brain stimulation decreases delta (1-4 Hz) while increasing beta and gamma (30-40 Hz) activity (Hlinka et al., 2010; Keeser et al., 2011). Furthermore, anodal prefrontal tDCS was shown to decrease and increase delta and beta power, respectively (Hlinka et al., 2010). Thus, the results obtained with regard to theta decrease and beta increase may support the presence of a more alert state (Barry et al., 2009) associated with multi-session tDCS.
Further, significant alpha increase at F3 in session eight can be confirmed by the alpha power increase during episodic and semantic memory tasks being associated with enhanced cognitive processing (Klimesch et al., 1994). Although the resting state qEEG was analyzed in this study, it can be hypothesised that multisession tDCS may have resulted in a more alert state being detected during the performance of cognitive tasks. Yet another possible explanation is that qEEG was recorded after tDCS combined with L2 instruction, which acts to reinforce the probability of semantic or episodic linguistic processing even during the resting state.
Significant increase in language network CSD z-score can possibly be confirmed by increased current density of prefrontal areas due to DLPFC anodal tDCS (Keeser et al., 2011). Moreover, these results are concordant with resting state fMRI scans that reveal increased connectivity of Broca's area and language related regions due to 1 mA, 17 minute stimulation in that location (Meinzer et al., 2012). Thus, the obtained results corroborate studies that report increased functional connectivity (Hlinka et al., 2010) and cortical/metabolic changes, not only in stimulated cortex but in broadly connected areas (Ardolino et al., 2005; Baudewig et al., 2001; Cho and Strafella, 2009; Lang et al., 2005). The question of how brain stimulation results in lasting changes in cortical excitability has recently been discussed (Keller et al., 2018). In agreement with (Keller et al., 2018), the findings reported here demonstrate an association between electrical stimulation and the excitability changes (i.e. CSD gain) in the language network. In other words, as for the above report, these findings corroborated that the observed changes in excitability were mostly dominant in regions anatomically and functionally close to the stimulated sites. Though there was a noticeable effect in the mean CSD of language network centers, this seemed to remain sustainable following eight sessions of subsequent tDCS intervention. Future studies may indicate whether a lower number of stimulation sessions would result in a comparable efficacy. Increased CSD within ROI in resting state corresponds to an increase in beta power. This would putatively be attributed to changes in neural plasticity, hence a possible increase in linguistic functional capacity and language learning.
This study was subject to some limitations. The sample size of the two groups should be increased as there were only eight subjects in each group. Longitudinal studies with iterative tasks over time as well as follow-up evaluation (qEEG in this case) tend to reduce subject participation. As such, the number of cases with full compliance to the experimental protocol only maintained eight subjects for each group (i.e. sham and tDCS study groups). This sample size for each group was determined by similar earlier studies and according to the sample size calculation of the number of subjects calculated to be required to achieve a power of 95% and significance level of 0.05. It should be noted that each subject underwent eight sessions of tDCS intervention including L2 instruction concurrent with tDCS. Therefore, for a total of 16 subjects, 128 tDCS intervention sessions and 32 qEEG acquisitions were necessary. Nevertheless, the linguistic and other cognitive effects (memory, reasoning, attention) of this protocol have been shown here to be suited to further research in which linguistic taskpositive qEEG and fMRI experimental designs may further inform more about tDCS-induced cortical language networks.
This study reports that multi-session anodal stimulation of Broca's area may modulate brain electrical activity and result in increased alertness during the resting state. This conclusion is supported by qEEG results that show decreased theta and increased beta activity in a large-scale language network. Changes in electrical activity and current density of language-related ROI may confirm the potential for tDCS to increase functional connectivity and affect subcortical areas. Further, the reported enhancement of cognitive-verbal performance due to brain stimulation may be supported by cortical and electrical changes of qEEG.
This research was approved by the ethical review board at Shiraz University of Medical Sciences, code 97901-1089. All participants gave written consent after being informed about the experimental procedures.
ZKS, AMK and MN conceived of the presented idea. ZKS, MS, SSY developed the theory and performed the computations. ZKS, MS, MN verified the analytical investigation and MN supervised the findings of this work. All authors discussed the results and contributed to the final draft of the present manuscript. ZKS and SSY carried out the experiments. ZKS and MN wrote the manuscript with support from SSY, MS, AMK.
The work was financially supported by DANA Brain Health Institute (DBHI), Shiraz, Iran.
Authors appreciate support from The Department of Neuroscience at Shiraz University of Medical Sciences, Department of foreign Languages and Linguistics at Shiraz University as well as, DANA Brain Health Institute (DBHI), Shiraz, Iran.
The authors declare no conflicts of interest statement.