

 , Bekim Gashi 2, Alessandro Scuotto 1, Glenda Cappello 1, Massimo Cocchi 1, Fabio Gabrielli 1, Jack A. Tuszynski 3
, Bekim Gashi 2, Alessandro Scuotto 1, Glenda Cappello 1, Massimo Cocchi 1, Fabio Gabrielli 1, Jack A. Tuszynski 31 LUDES Foundation, Smart City, Kalkara, 1001, Malta
2 Department of Biology, University of Prishtina "Hasan Prishtina", Prishtina, 10000, Kosovo
3 Department of Physics, University of Alberta, Edmonton, Alberta, T6G 2J1, Canada
Abstract
Living organisms tend to find viable strategies under ambient conditions that optimize their search for, and utilization of, life-sustaining resources. For plants, a leading role in this process is performed by auxin, a plant hormone that drives morphological development, dynamics, and movement to optimize the absorption of light (through branches and leaves) and chemical "food" (through roots). Similarly to auxin in plants, serotonin seems to play an important role in higher animals, especially humans. Here, it is proposed that morphological and functional similarities between (i) plant leaves and the animal/human brain and (ii) plant roots and the animal/human gastro-intestinal tract have general features in common. Plants interact with light and use it for biological energy, whereas, neurons in the central nervous system seem to interact with biophotons and use them for proper brain function. Further, as auxin drives the "arborescence" of roots within the soil, similarly serotonin seems to facilitate enteric nervous system connectivity within the human gastrointestinal tract. This auxin/serotonin parallel suggests the root- branch axis in plants may be an evolutionary precursor to the gastrointestinal-brain axis in humans. Finally, it is hypothesized that light might be an important factor, both in gastrointestinal dynamics and brain function. Such a comparison may indicate a key role for the interaction of light and serotonin in neuronal physiology (possibly in both the central nervous system and the enteric nervous system), and according to recent work, mind and consciousness.
Keywords
- Gastrointestinal-brain axis
- tryptophan
- serotonin
- auxin
- light
- quantum brain model
- biophotons
- plants
According to Darwin's theory of evolution by natural selection, all organisms exhibit a tendency under ambient conditions to optimize their means of access to, and utilization of, life-sustaining resources. Considerable empirical evidence reveals that this strategy involves nutrients and other energy-giving resources (water, light, heat, shelter) as metabolism is a necessary pre-condition for life, but requires various energy-rich inputs and components for enzymatic reactions to enable such efficient utilization. As demonstrated in plants, an important leading role in pursuing such aims is played by auxin, which is a hormone produced in the stem tip that promotes cell elongation. Auxin moves to the darker side of the plant, causing the cells there to grow larger than the corresponding cells on the lighter side. It acts to drive plant growth, development of morphology, dynamics, and even their movement in order to optimize the absorption of an essential vital element: light (through branches and leaves), and "food" in the form of proper chemical substances (absorbed through roots). Comparable features and behaviors can be identified to provide parallels between plants and humans that suggest evolutionary advance inherited from ancient ancestors, retained and adapted for the needs of the human body. In particular, as does auxin in plants, serotonin seems to play a similar role in higher animals, especially humans [1,2]. Serotonin is a monoamine neurotransmitter bio-chemically derived from tryptopan, which is predominantly found in the central nervous system (CNS), the gastrointestinal (GI) tract and the blood platelets of animals, including humans. Recent findings from different lines of research newly suggest the possibility of common strategies developed by both plants and humans that may be understood through their shared remote ancestry. In particular, morphological and functional similarities between plant leaves and animal/human brain, as well as plant roots and animal/human GI can be outlined in terms of general morphological and functional features.
Pertinent to this connection, the following questions are proposed: given a parallel between serotonin in animals and auxin in plants, could light and food be factors playing similar roles in plants and humans, respectively? Is it plausible that the root-leaf axis is the evolutionary ancestor of the GI-brain axis? The latter has been the subject of keen research interest in recent years. However, to the best of the authors knowledge, virtually no attention has been given to a possible root-leaf axis in plants. In a recent paper, Mender [3] drew attention to the parallel between quantum phenomena involved in plant and algae photosynthetic processes on the one hand and consciousness in the human brain on the other hand. He noted that energy harvesting by photosynthesis in plants could be viewed as a precursor to complex quantum process powering neural correlates of human consciousness. In the present paper a direct parallel between the specific mechanisms involving the root-leaf axis in plants and the GI-brain axis in humans is elaborated. With such a parallel, the reader's attention is drawn to this idea which here is elaborated as a hypothesis that possibly concerns such a relatable array of conjectures.
One of the fundamental aspects of plant survival is its light absorption capability, since light is a primary source of living energy on our planet driving the photosynthesis that propels plant growth. Plants then become sources of food for higher organisms in a food chain that eventually terminates with the human consumption of both plants and animals for survival. Light has another fundamental role in the Plant Kingdom: it not only provides energy for photosynthesis but also transmits reliable information, mainly regarding local environmental conditions. Light intensity, and the direction it comes from, informs a plant that absorbs it about its immediate environment. Light may blocked or its intensity reduced depending upon the distribution and density of organic and inorganic materials surrounding a given plant. Recently, it has been convincingly and realistically hypothesized that plants could have developed a form of primitive vision ability allowing them to receive and process large amounts of information provided by light [4, 5]. Furthermore, light interaction with auxin, elicits local responses as well as long-distance signalling. Light coordinates growth between the shoot and the root and affects plants from roots to leaves by modulating their growth and development [6,7]. To expand on this Halliday et al. [7]is quoted: "Light imposes a strong influence on multiple facets of the auxin system, controlling auxin levels, transport, and responsiveness}". In particular, "light triggers auxin synthesis in developing young leaves… whereas auxin transported to the root is required for lateral root emergence and primary root elongation". Supplementary to the above references, there is strong light-auxin interaction in terms of light intensity affecting auxin levels, which is turn coordinates development between the shoot and the roots, manipulating plant growth in order to tune and adapt a plant to its light environment. Such interactions appear to be the basis of plant plasticity [8].
It is apparent that the dynamics generated by light-auxin interactions seems to involve phytochrome. For example, in Arabidopsis, the mechanism may be linked to phytochrome A and B. These enzymes can impose a strong influence on auxin levels in plants, by acting through tryptophan metabolism enzymes (SUR2 and TAA1) and, as a feedback mechanism working on available auxin, transcriptionally regulating several enzymes encoding genes (GH3 family) as reported by Franklin and Whitelam [9] and Halliday et al. [7]. Further, the role of light is involved with both the distribution of auxin from shoot to root and its flux. Light pathways may modify auxin distribution by controlling the abundance of auxin efflux carriers such as PIN-FORMED proteins and p-glycoproteins of the ATP-binding cassette family B (ABCB) transporter molecules, through phytochromes and, probably also, cryptochromes.
Furthermore, light moderates sensitivity to auxin within the cell by imposing control on the nuclear auxin response pathway. It can dampen or amplify the response to auxin and thus regulate signal transduction such that light can target responses at specific locations in the plant.
Interestingly, light and auxin seem to have common gene targets. In fact, at the molecular level, auxin activates the transcription of three gene families: Aux/IAA, SAUR, and GH3. Several studies [7,9,10,11,12,13,14] suggest that light (through phytochrome) seems to be involved in regulating the expression of several of the same genes belonging to the gene families: Aux/IAA, SAUR, and GH3. There are a number of transcription factor convergence points that enable the coordinated regulation of genes by light and auxin.
Auxin is the main factor involved in the optimization of light capture by leaves [6,7], thus it can play out different strategies such as directly driving plant growth, its morphology, and dynamics, from shape, via branch structure and arborescence, to the arrangement of leaves, and even their "movement". For instance, the turning of the leaf toward the direction of light, its source of energy, depends upon the rearrangement of the cytoskeleton inside the cells of leaves through a mechanism directly involving auxin [1,6,7].The actions of auxin in plants strongly resembles the well-known actions of serotonin in neuronal development and adult neuroplasticity in mammals [15,16]. Strong evidence indicates that serotonin performs a major driving role in neuronal morphology and dynamics [17,18,19].
Accordingly, comparable to the way in which auxin strongly affects the structure of branches and leaves (to optimize photosynthesis oriented light absorption) so in animals/humans, serotonin strongly influences the morphology of neurons involved in CNS neuronal networking. Moreover, auxin and serotonin have biochemical structural properties in common and hence exhibit some analogous biological functions. Biochemically, they are both tryptophan metabolites, they have a very similar molecular structure and hence similar chemical and physical properties as argued above and in the references cited there. Unsurprisingly, this strongly suggests that auxins perform similar functions in plants and animals. Therefore, the following questions arise: In analogy to auxin in plants, is it possible that there is a type of “light interaction” in the CNS, involving serotonin and neuronal development and morphology? Is there a neuronal equivalent to arborescence, in analogy to that exhibited by the branches and leaves of plants? Would this mechanism involve neurons interacting with light or even actively searching for light or doing something similar? Are there any other possible similarities and what might their roles be?
A growing body of evidence supports the idea of the involvement of biophotons within the inner workings of the human brain. Several possible ways of interaction with neurons have been suggested and they indicate that photons may play a key role in neural information processing, electromagnetic signalling, and may be involved in quantum mechanical brain mechanisms, as elaborated on in several recent reports [20,21,22,23]. In particular, mitochondria and microtubules in neurons as well as myelinated axons have been hypothesized to serve as photonic waveguides [24,25,26,27]. Also, it has been shown that glutamate (which is by far the most abundant neurotransmitter molecule in the nervous systems of vertebrates—on the basis of its involvement in over 90% of synaptic connections in the human brain) could induce biophotonic activities and transmission in neural circuits [28].
Hence, in human brain, neurons seem to interact with biophotons, which are light quanta spanning the near-UV to near-IR frequency range. In an analogous manner to plants, which interact with light and use it for biological energy, it is feasible that neurons are able to interact with photons as explained in detail by Kumar et al. [24]. While as of yet there are no experimentally documented specific functions of biophotons in the brain, the speed and precise nature of photon-receptor interactions offer an attractive mechanism that evolutionary exploration could have found of use for in optimizing brain function, for example, in the coordination and synchronization of cognitive functions. Future studies should be directed toward addressing specific roles for biophotons in the brain. It is reasonable to expect that a biological organ such as the brain, which operates at great efficiency using electrical signals involving action potentials, also uses electromagnetic signals that could provide an independent channel of communication [29]. A double comparison between plant leaves and animal neurons can be summed up as follows: they both use auxin/serotonin, they both interact with light. Is this an accidental coincidence or is this correlation significant from an evolutionary perspective?
It has even been suggested that interactions of neurons with biophotons could be a fundamental element involved in the emergence of cognitive abilities or even the basis for consciousness [2,30]. Therefore, since the auxin interaction with light is a driving force in plants providing a mechanism for the optimization of biological energy absorption and utilization, serotonin and biophotons could drive neurons in order to organize an active "mind" or be a spark that ignites "consciousness". This is, of course, a bold and unproven hypothesis with a somewhat provocative intent, which we hope will generate a robust debate in the research community.
The fundamental element for plant survival is their "food" intake, represented by water, nitrogen and important minerals and micronutrient compounds searched for and captured by roots. Plants can send their roots deep into the soil to capture what is needed and, as in the case of leaves, above ground to search for optimal light exposure. Movement performed underground by roots depends upon auxin [31]. Thus, auxin controls and steers root growth, plasticity, and movement so as to optimize "food" search and absorption.
Is there a similar behavior applicable to animal/human physiology, possibly driven once again by serotonin? What about the GI? Serotonin's role in the GI has been very well studied. Among several suggested aims and functions involving it, a most interesting one is linked to the Enteric Nervous System (ENS), a neural network located in the GI. In fact, serotonin has been observed to stimulate stem cells to divide and give rise to new neurons of the ENS even in adult animals [32]. Its ability appears to be essential for post-natal and adult health. Enteric neuronal serotonin affects growth/maintenance of the ENS in terms of neurogenesis (as well as intestinal mucosa) [33,34]. Moreover, the ability of serotoninergic neurons to sculpt the ENS potentially enables environmental stimuli that alter the activity of serotoninergic neurons to produce long-lasting changes in the structure and function of the ENS [35]. Hence, as auxin drives "arborescence" of the roots for plants inside the soil, so too does serotonin appear to lead the wiring of ENS neurons in the human GI?
It is well known that the action of auxin on plant roots is clearly oriented toward the search for and intake of food. These processes include information transmission regarding environmental conditions and this process is hypothesized to involve feedback loops for back-and-forth information transmission. What is known about ENS function with analogous properties to those of the auxin-root system? The ENS is exposed to, and interacts with, the outer (microbiota, metabolites, and nutrients) and inner (immune cells and stromal cells) microenvironment of the GI. Moreover, two types of enteric neuron send signals: Some of which densely innervate intestinal villi and detect food, while others target stomach and intestinal muscle and sense stretch [36,37].
A clear comparison between plant roots and the animal (human in particular) GI (ENS) seems to involve the following properties. They both use auxin/serotonin for their organization and functionality as well as for information transmission. Also, they both work in order to optimize food intake and absorption interacting with substance of the outside world in a similar way. Moreover, they both need the presence of symbiotic bacteria for their performance: these bacteria belong, above all, to philum Firmicutes, particularly to the class Clostridida [38,39]. Then again from the morphological point of view, the spatial geometry of roots and the GI seems to be similar. In particular, it is easy to recognize fractal structures in the roots of plants [40] as well as in the animal/human GI, the ENS in particular [41]. From this perspective, at least for some elements, the ENS clearly mimics and directly resembles the role, function, and dynamics of the corresponding properties of plant roots.
Interestingly, auxin is linked to leaves and roots in plants just as, in humans; serotonin is linked with the brain neurons in the CNS and with the GI neurons in the ENS, respectively. Given this connection the peculiar role of the neuron as a serotonin-coupled cell should be stressed in the plant-human comparison. Notably, the morphology of roots resemble GI morphology, from the fractal perspective, ENS neurons in particular. Interestingly, also leaves, branches, and canopies follow fractal geometry development [42] as well as brain neuron arborescence [43]. It is well known that such a geometry is widespread in nature to have maximize capillary penetration, provide wide diffusion, and optimize exchange surfaces (e.g. think of the structure of the lung) [41,44,45].
Azmitia [1] claims that one of the serotonin (and similar compounds') roles in plants and humans seems to be driving cells "in order to tract the source of relevant stimuli". As mentioned above, the role of auxin in plants is its interaction with light in the case of leaves and the search for food in the case of roots. In the same way, the role of serotonin in the human body could be hypothesized to interact with light (in the form of internally generated biophotons) in the brain and to search for food in the GI. Consequently, the auxin/serotonin parallel suggests a comparison between what could be somehow called the root-branches axis in plants as the ancient precursors of the GI-brain axis (GBA) in humans (see Fig.1).
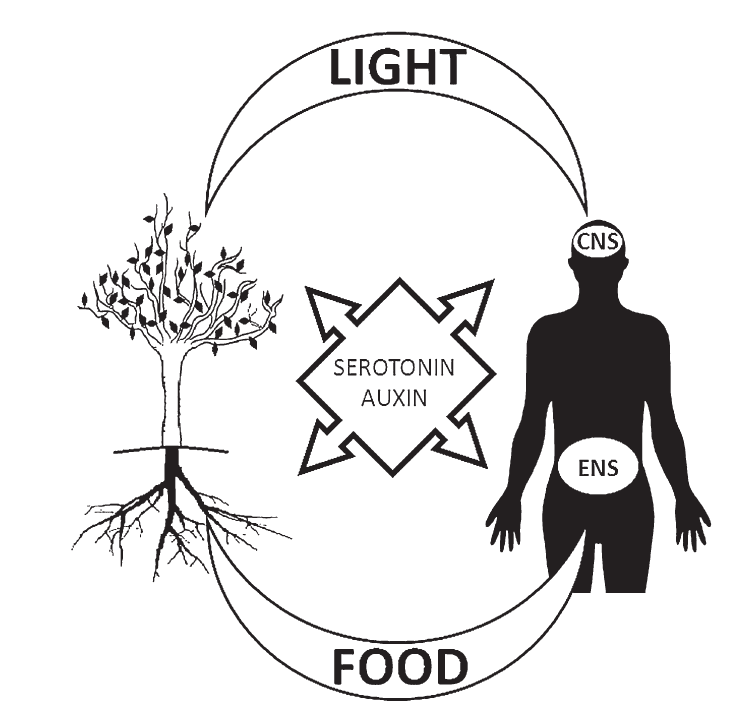 Fig. 1.
Fig. 1.An illustration of the roles of auxin in plants and serotonin in humans. Auxin is linked to leaves and roots in plants just as serotonin in humans is linked to brain neurons (CNS) and with GI neurons (ENS), respectively. Both auxin and serotonin interact with light, the former through branches and leaves, the latter through the CNS. On the other hand, auxin and serotonin interact with food, the former through roots, the latter through the ENS.
It should be noted that in humans, serotonin synthesis from tryptophan uses TPH (tryptophan hydroxylase) enzymes and two separate types of TPH are known: TPH1 and TPH2. The first is located in non-brain tissue and it is responsible for producing most of the serotonin found in the body, including the blood [46]. Alternatively, TPH2 is entirely restricted to neurons of the raphe nuclei and the ENS and is the enzyme responsible for producing all of the serotonin in the brain and in the ENS [47]. So, interestingly, the serotonin of the brain and of the ENS are both synthesized through a common enzyme, TPH2 (different from the serotonin available in the rest of the body available through the TPH1 synthesis), thus suggesting, once again, a possible common root. In other words, the GBA could be seen as an evolution of the dynamics of branches (leaves) and roots because of the similar aim and function of the common family of compounds, the tryptophan metabolites, i.e. serotonin and auxin.
Hence, in terms of the retention and refinement of evolutionary advances, the hypothesis is based on the lines of reasoning presented above, suggesting that the human GI could be seen as playing an analogous role to that of the root of a plant. The evolutionary step that comprised the acquisition of great movement ability (e.g. crawling, walking, and running) gained by animals in general and humans in particular, also necessitated a means for making access to their food sources "portable". Animals had somehow to take with them a "piece" of the earth in order to find vital nutrition elements "on the go." Therefore, evolution may have led to the generation of a food container (the gastro-intestinal tract) that can be carried along with the now mobile living systems (animals).
From this perspective, what could be the evolutionary meaning of brain neurons in humans? They have been compared to plant leaves but branches and leaves are in the open air, neurons are not. In humans, such "open air" has been enclosed in the skull. Therefore, the head of an animal can be viewed within the same evolutionary point of view as a parallel to the open environment of a plant. However, the skull confines and isolates the animal's brain and prevents most of the externally generated photons from reaching the brain's neurons. Nonetheless, the parallel between neurons and leaves is not entirely lost due to the fact that the skull encloses the brain. While the source of light for a plant is the sun, the source of biophotons is not external, but produced within brain cells as a byproduct of metabolism. In fact, possible sources of biophotons have been identified as the result of mitochondrial respiration [48, 49] or lipid oxidation [50,51]. Nevertheless, miniature photon-producing version of the sun may be present within us. In this sense, our head may be interpreted as a luminous environment, made "portable", and evolved to become self-illuminated.
The proposed scenario reported here suggests light as the main factor affecting the function of leaves in plants in terms of capturing sunlight and storing its energy, and equally an important factor for human neurons in terms of biophotons and their utilization in cognitive processes and possibly playing a crucial role in consciousness.
At the same time, it has been well known for decades that light influences the growth and development of roots [52,53,54]. In a recent paper, Lee et al. [55] show that in plants, Arabidopsis thaliana in particular, "light also influences root growth and development". Specifically, they demonstrated that "light was efficiently conducted through the stems to the roots", and that "underground roots directly sense stem-piped light to monitor the aboveground light environment".
In view of the above arguments, if the human CNS is similar to plant leaves in terms of light behavior, and if neurons of the CNS and neurons of the ENS are similar with the ENS being similar to plants root, then it seems reasonable that the ENS should be involved in light-oriented dynamics. Strictly speaking, this hypothesis suggests that light could be an important factor in the brain and in GI dynamics, whose interactions may play a role similar to that played by the root-leaf axis in plants, i.e. as a messenger of the "environmental light" condition. Thus, according to what is proposed above, in humans the "environmental light" is represented by brain biophotons so, a possible hypothesis is that "human light" could be a messenger within the GI-brain axis. It is our contention that at the very least biophoton involvement in the ENS should be seriously considered.
It has been described that auxin plays a fundamental role in plants, from roots to leaves. At the same time, it has been shown that a parallel role seems to be played by serotonin in humans, from the GI to brain. In order to perform its tasks, auxin interacts with light. On the other hand, in humans, serotonin seems to be involved in neuron dynamics, development and behavior but it still remains unknown as to what its possible interaction with light mat be (beyond its known involvement in the activation of enzymatic conversion of 5-hydroxytryptophan to serotonin [56] and in other physiological processes [57]). In other words, we have argued that in humans serotonin and light are involved in similar cellular processes, which could be used for the same purposes and could be playing similar roles. However, in the absence of the crucial experimental demonstration of biophoton generation and utilization in the brain, what specific mechanisms are involved in these interactions still remains to be determined.
In an earlier article it was proposed a possible quantum interaction may be based on the presence of an indole ring within the serotonin molecule Tonello et al. [2] but the suggested mechanism, while feasible, remains no more than conjecture. Nonetheless, the plant/human comparison seems to strongly suggest the key role of the interaction between light and serotonin as a fundamental element for insight into neuron physiology (maybe both in the CNS and ENS). Such an understanding may lead to the unlocking of the mysteries of the mind and consciousness. It is worth noting that "Fluorescent and absorbing substances should interfere with such a biophoton communication system. Of all natural amino acids, nature has chosen the aromatic ones with the strongest fluorescence, tryptophan, phenylalanine and tyrosine as precursors for the neurotransmitters involved in mood reactions: serotonin, dopamine and norepinephrine" [23].
Unfortunately, in plants, things are not as clear. Halliday et al. [7] claim that "Although researchers have uncovered many examples of light and auxin signal integration, the future challenge is now to generate a model of the light-auxin network, with spatial and temporal resolution, that can predict plant behavior in response to environmental light stimuli". It appears as a final conjecture that a better understanding of the auxin-light interaction could also lead to a better understanding of the cognitive mechanisms taking place in the brain by opening a new line of investigations focused on the electromagnetic interactions involving biomolecules such as auxin, in addition to the well-studied electric signalling via action potentials and chemical signalling via neurotransmitters. The subtleness of electromagnetic interactions and their precise coupling to molecular absorption due to resonance effects offer numerous advantages and could explain a number of enigmatic features of the human mind [58].
LT thanks Prof. Claudio Morellato for his lifelong priceless teachings. JAT thanks NSERC (Canada) for funding his research.
All authors declare no conflicts of interest.