



 , Katarzyna Skrypnik 1, Paweł Kołodziejski 3, Joanna Suliburska 1,*
, Katarzyna Skrypnik 1, Paweł Kołodziejski 3, Joanna Suliburska 1,*1 Department of Human Nutrition and Dietetics, Faculty of Food Science and Nutrition, Poznan University of Life Sciences, 60-624 Poznan, Poland
2 Research Organization for Agricultural and Food, National Research and Innovation Agency, 16911 Bogor, Indonesia
3 Department of Animal Physiology, Biochemistry and Biostructure, Faculty of Veterinary Medicine and Animal Science, Poznan University of Life Sciences, 60-637 Poznan, Poland
Abstract
Phosphorus is an essential component of bone and energy metabolism; however, the mechanism involved in the regulation of phosphorus under postmenopausal conditions remains insufficiently understood. Interestingly, the disruption of estrogen status can alter mineral homeostasis. Meanwhile, phytoestrogens and probiotics may offer a non-pharmacological strategy to support mineral homeostasis; nonetheless, evidence of their impact on phosphorus distribution remains limited. Thus, this study aimed to evaluate the effect of daidzein, tempeh, and Lactobacillus acidophilus on phosphorus distribution in ovariectomized (OVX) rats.
A total of 64 female Wistar rats were allocated to sham-operated (n = 8) or OVX (n = 56) groups. Following a three-week calcium-deficient diet, the OVX rats were randomized to seven dietary interventions for six weeks: control (O), alendronate (OB), daidzein (10 mg/kg diet/day; OD), tempeh (250 g/kg diet/day; OT), probiotic L. acidophilus (1 × 1010 CFU/day; OL), daidzein + probiotic (ODL), or tempeh + probiotic (OTL). Food intake, body mass, and phosphorus concentrations in feces, the femur, kidney, pancreas, spleen, heart, and liver were analyzed.
OVX groups (O, 366 ± 14 g; p = 0.01; OD, 361 ± 18 g; p = 0.02; OL, 358 ± 29 g; p = 0.04) gained significantly more body mass than sham controls (311 ± 30 g). Dietary interventions with daidzein, tempeh, and L. acidophilus enhanced phosphorus deposition in the femur, kidney, and pancreas compared with the OVX controls. The OT group showed the highest femoral phosphorus levels (178,111 ± 32,628 mg/kg d.w., p = 0.02), while the ODL group revealed elevated phosphorus levels in the kidney (12,966 ± 462 mg/kg d.w.; p = 0.01). The OD (914 ± 115 mg/kg d.w.; p < 0.001), ODL (913 ± 39 mg/kg d.w.; p < 0.001), and OTL (926 ± 70 mg/kg d.w.; p < 0.001) groups showed significantly increased levels of pancreatic phosphorus. A positive correlation was noted between the pancreatic and femoral phosphorus levels (r = 0.33; p = 0.02). Alendronate treatment had no significant effect.
Dietary daidzein, tempeh, and probiotics modulate phosphorus distribution in postmenopausal rats. These results support the translational potential of dietary bioactives to improve phosphorus metabolism in postmenopausal women.
Keywords
- minerals metabolism
- phytoestrogens
- probiotics
- postmenopausal
- women’s health
Menopause is characterized by a marked decline in circulating estrogen, profoundly affecting mineral metabolism and skeletal health [1]. Estrogen deficiency disrupts the balance between bone formation and resorption, leading to increased bone turnover, reduced bone mineral density, and heightened susceptibility to osteoporosis and fractures [2]. While calcium homeostasis has been extensively investigated [3], less is known about the regulation of phosphorus, a critical mineral for bone structure and systemic metabolic processes. Phosphorus contributes to hydroxyapatite formation in bone and participates in adenosine triphosphate (ATP) synthesis, enzymatic reactions, and intracellular signaling [4]. Its systemic levels are tightly controlled through intestinal absorption, renal excretion, and hormonal regulation, primarily via fibroblast growth factor 23 (FGF23), parathyroid hormone (PTH), and vitamin D [5]. Despite its physiological importance, phosphorus metabolism under postmenopausal conditions remains poorly characterized, particularly in relation to dietary interventions targeting mineral homeostasis. Previous studies, including our work on probiotic and isoflavone combinations, have demonstrated significant effects on calcium status and bone metabolism biomarkers [6, 7]; however, tissue phosphorus deposition and its regulation under estrogen-deficient conditions have received little attention.
Dietary interventions offer a promising strategy to modulate mineral metabolism in postmenopausal populations. Isoflavones, particularly daidzein, are phytoestrogens abundant in soy and fermented soy products such as tempeh [8]. Daidzein exists as both glycoside and aglycone forms, with the latter exhibiting higher bioavailability [9]. Preclinical and clinical studies indicate that daidzein can modulate bone turnover and enhance calcium absorption, partially compensating for estrogen deficiency [10, 11]. However, its direct influence on phosphorus homeostasis has not been systematically investigated. Fermented soy products, including tempeh, offer additional benefits: fermentation enhances isoflavone bioavailability, generates bioactive peptides, and improves mineral absorption by reducing antinutritional factors such as phytates [8]. These properties may facilitate phosphorus absorption and systemic utilization, supporting skeletal and metabolic health in estrogen-deficient states. Similarly, probiotics such as Lactobacillus acidophilus can modulate gut microbiota composition and activity, liberating minerals from dietary complexes through enzymatic processes such as phytase-mediated hydrolysis [12, 13]. Despite these mechanistic insights, empirical data on the effects of isoflavones and probiotics on phosphorus distribution in postmenopausal models remain scarce.
Collectively, while calcium metabolism has been extensively explored, phosphorus homeostasis in postmenopausal conditions is understudied [14], and its modulation by dietary bioactives remains largely unknown. In addition, direct comparisons of dietary strategies with pharmacological interventions, such as alendronate, which inhibit osteoclast-mediated bone resorption, are lacking. Addressing these gaps is crucial for understanding how diet-based interventions may influence systemic phosphorus regulation in estrogen-deficient states.
The present study aimed to systematically evaluate the effects of three dietary interventions—pure daidzein, fermented soy product (tempeh), and probiotic L. acidophilus—on phosphorus homeostasis in ovariectomized rats as a model of postmenopausal bone loss. These interventions were chosen to represent distinct but complementary dietary strategies: daidzein as a phytoestrogen influencing mineral metabolism, tempeh as a naturally fermented isoflavone source, and L. acidophilus as a probiotic potentially enhancing nutrient bioavailability. It was hypothesized that daidzein and tempeh enhance phosphorus deposition in bone and metabolically active organs, reflecting improved mineral bioavailability and systemic distribution, while probiotics link to phosphorus availability through gut microbiota-mediated mechanisms. Furthermore, combinations of daidzein, tempeh, and probiotics may produce synergistic effects, and these patterns of phosphorus modulation were anticipated to differ from those observed with alendronate treatment. This investigation provided a rationale for exploring both single-component and combined dietary strategies to address critical knowledge gaps in postmenopausal phosphorus metabolism, with potential translational relevance for nutritional approaches to skeletal and metabolic health.
Three-month-old female Wistar rats were selected as the animal model and obtained from the Nencki Institute of Experimental Biology (Warsaw, Poland). Animals were maintained on a purified diet based on the AIN-93M formulation, supplied by Zoolab (Sędziszów, Poland). Wistar rats were selected due to their established sensitivity to estrogen deficiency and their reproducibility as a model for postmenopausal metabolic alterations [15]. The primary dietary ingredients consisted of soybeans of the Augusta cultivar, sourced from the Poznań University of Life Sciences (Poznań, Poland), while purified daidzein was provided by Gentaur Molecular Products BVBA (Kampenhout, Belgium). Calcium citrate tetrahydrate, incorporated as the mineral source in the basal diet, was purchased from Warchem Sp. z o.o. (Warsaw, Poland). Tempeh flour was prepared using Rhizopus oligosporus NRRL 2710, a strain obtained from the Agricultural Research Service Culture Collection (Peoria, IL, USA) [16]. The probiotic strain Lactobacillus acidophilus DSM 20079 was supplied by the Leibniz Institute DSMZ (Braunschweig, Germany) and prepared according to a method described in a previous study [17].
The rats were housed in pairs in stainless steel cages with enamel-coated bases,
equipped with wire-mesh tops to allow adequate ventilation. Each cage contained a
polycarbonate drinking bottle, regularly refilled with fresh water, and
stainless-steel feeders to provide consistent access to experimental diets. The
choice of stainless-steel cages minimized contamination risks and facilitated
routine cleaning and disinfection, ensuring a hygienic and controlled
environment. Environmental parameters were strictly regulated, with the animal
room maintained at 21
A total of 64 female Wistar rats were included in this study. After a one-week acclimatization period, the animals were randomly assigned into two experimental groups: a sham-operated control group (n = 8) and an ovariectomized (OVX) group (n = 56). Bilateral ovariectomy was performed in the OVX group under general anesthesia using a ketamine/medetomidine combination (Cepetor KH, CP-Pharma Handelsgesellschaft mbH, Burgdorf, Germany), administered by an experienced veterinary surgeon under aseptic conditions. The sham-operated animals underwent identical surgical procedures, with the exception that the ovaries were left intact. Immediately following surgery, all rats were placed on heated recovery pads maintained at 30 °C and continuously observed until full consciousness was regained. Postoperative monitoring was performed daily for seven consecutive days, including assessments of general behavior, surgical wound integrity, food and water intake, and body weight. Analgesic support was provided when required, and no adverse events or losses occurred during this period. Throughout the recovery phase, all animals received a semi-synthetic AIN-93M diet and had unrestricted access to fresh drinking water. This recovery protocol was designed to ensure animal welfare, minimize postoperative stress, and provide stable baseline conditions before dietary interventions.
After a one-week postoperative recovery period, the rats were weighed, and the
ovariectomized cohort was randomly assigned to experimental subgroups to ensure
comparable baseline body weights across groups. Randomization was conducted to
establish similar weight distributions at the start of the dietary intervention.
Animals were randomly assigned to treatment groups by an independent researcher
not involved in outcome assessment. Due to the laboratory setting and fixed
biochemical procedures, blinding during assay measurement was not feasible, but
sample handling followed a standardized protocol to minimize bias. To standardize
baseline mineral status and simulate conditions that exacerbate post-menopausal
bone loss, a calcium-deficient diet was administered for three weeks before the
main dietary interventions. This short-term calcium restriction has been used
previously to induce a reproducible state of reduced calcium availability and was
selected based on published evidence indicating that three weeks is sufficient to
produce measurable calcium depletion in rodents [18]. During this period, food
intake was recorded at the cage level and body weights were measured weekly to
monitor tolerance and ensure animal welfare. The basal diet contained calcium at
5.06 g/kg [7] and phosphorus at 1.10 g/kg, yielding a calcium-to-phosphorus ratio
of approximately 4.6:1 on a mass basis (
After the three-week calcium-deficient induction period, all animals were
reassigned to their respective experimental diets for a six-week intervention
phase. The sham-operated group (S; n = 8) was returned to the standard AIN-93M
maintenance diet containing adequate calcium, serving as the physiological
reference. The ovariectomized rats (n = 56) were then randomized into seven
dietary intervention groups (n = 8 per group), with randomization stratified by
baseline body weight to minimize allocation bias. The experimental groups were as
follows: ovariectomized controls (O, n = 8) received the standard AIN-93M diet;
alendronate treatment (OB, n = 8) consisted of AIN-93M supplemented with a
therapeutic dose of alendronate; daidzein supplementation (OD, n = 8) received
AIN-93M enriched with daidzein at 10 mg/kg diet; tempeh supplementation (OT, n =
8) received AIN-93M containing tempeh flour at 250 g/kg diet; probiotic treatment
(OL, n = 8) received AIN-93M with a daily administration of L.
acidophilus at 1
| Group | Number of rats | Description | Composition | Dosage |
| S | 8 | Sham | AIN-93M | - |
| O | 8 | OVX | AIN-93M | - |
| OB | 8 | OVX | AIN-93M + alendronate | 3 mg/kg body weight |
| OD | 8 | OVX | AIN-93M + daidzein | 10 mg/kg of diet |
| OT | 8 | OVX | AIN-93M + tempeh flour | 250 g/kg of diet |
| OL | 8 | OVX | AIN-93M + L. acidophilus | 1 × 1010 CFU/day |
| ODL | 8 | OVX | AIN-93M + daidzein + L. acidophilus | 10 mg/kg of diet + 1 × 1010 CFU/day |
| OTL | 8 | OVX | AIN-93M + tempeh flour + L. acidophilus | 250 g/kg of diet + 1 × 1010 CFU/day |
S, Sham group receiving AIN-93M; O, Ovariectomized (OVX) group receiving AIN-93M; OB, OVX group receiving AIN-93M with alendronate; OD, OVX group receiving AIN-93M with daidzein; OT, OVX group receiving AIN-93M with tempeh; OL, OVX group receiving AIN-93M with probiotics; ODL, OVX group receiving AIN-93M with daidzein and probiotics; OTL, OVX group receiving AIN-93M with tempeh and probiotics.
The dosages of all interventions were selected based on established experimental practice and translational considerations. Alendronate was included as a pharmacological reference group at a dose of 3 mg/kg body weight, administered once weekly. This dose was chosen because alendronate is a well-characterized anti-resorptive agent for postmenopausal osteoporosis, and its efficacy has been consistently demonstrated in ovariectomized rat models, particularly with respect to bone remodeling and fracture healing [19]. To ensure consistent exposure, dosing was recorded for each animal in dosing logs maintained throughout the study. Standard AIN-93M diet served as the vehicle for the compound. Body weight was measured weekly to adjust the dose accordingly. No pharmacodynamic markers were assessed in this study; however, alendronate’s established effects on bone tissue serve as a reference for expected skeletal outcomes.
The doses of daidzein and tempeh were established according to the isoflavone profile quantified in the tempeh preparation applied in this experiment. Analytical assessment showed that 250 g of tempeh provided approximately 10 mg of daidzein equivalents. Based on this composition, the experimental diet was formulated to contain 250 g/kg of tempeh powder, which corresponds to an estimated 10 mg/kg of daidzein. For consistency across treatments, the diet supplemented with pure daidzein was adjusted to deliver the same concentration (10 mg/kg diet) in its aglycone form. This strategy allowed a controlled comparison between isoflavones supplied as conjugated glycosides in tempeh and as the isolated aglycone in pure form. Since glycoside conjugates require microbial hydrolysis to release bioactive aglycones during digestion [20], the distinction between these forms was an important consideration in evaluating bioavailability and biological effects. The chosen dosages are consistent with previously reported dietary intakes shown to benefit bone health in ovariectomized rodent models [21, 22].
For the probiotic group, L. acidophilus DSM 20079 was administered at 1
At the conclusion of the six-week dietary intervention, the rats underwent a 4–6 hour fasting period to minimize the influence of recent food intake on body weight and biochemical measurements. Animals were then humanely euthanized by decapitation, a rapid and widely accepted method that minimizes pain and distress. Immediately following euthanasia, relevant tissues and organs, including feces, spleen, femur, heart, liver, kidney, and pancreas, were collected for subsequent phosphorus analysis. Throughout the study, no deaths, exclusions, or adverse events occurred, and all experimental procedures were performed in compliance with national and institutional guidelines for animal care.
Phosphorus concentrations in experimental diets and feces samples were determined by dry ashing. Specifically, 2 g of diet and 1 g of feces were placed in porcelain crucibles and incinerated in a muffle furnace at 450 °C until complete mineralization was achieved. The resulting ash was dissolved in 1 N ultrapure nitric acid (Merck, Kenilworth, NJ, USA) for subsequent analysis.
For tissue samples—including feces (mg/kg), spleen (mg/kg), femur (mg/kg), heart (mg/kg), liver (mg/kg), kidney (mg/kg), pancreas (mg/kg)—approximately 0.5–1 g of each tissue was digested using 65% (w/w) ultrapure nitric acid (Merck, Kenilworth, NJ, USA) in a microwave digestion system (Speedwave Xpert, Berghof, Eningen, Germany). After digestion, the samples were diluted appropriately with deionized water to achieve phosphorus concentrations within the working range of the colorimetric assay.
Phosphorus quantification was performed using the ammonium molybdate–vanadate
colorimetric method, as described previously by Zubek et al. (2015)
[25], with minor modifications. In brief, the molybdenum-vanadate reagent was
prepared by mixing an ammonium heptamolybdate tetrahydrate (Sigma‑Aldrich, St.
Louis, MO, USA) solution with ammonium metavanadate (Sigma‑Aldrich, St. Louis,
MO, USA) in nitric acid, and then diluting the mixture to volume with deionized
water. Standard phosphorus solutions (25–300 µg/mL) were prepared
from a 1 mg/mL KH2PO4 (Merck, Darmstadt, Germany) stock solution for
calibration purposes. For each sample, 2.5 mL of the prepared solution was mixed
with 2.5 mL of the molybdenum-vanadate reagent and incubated at room temperature
(~20 °C) for at least 10 minutes. Absorbance was
measured at 430 nm using a UV–Vis spectrophotometer (Nanocolor UV/VIS,
Macherey-Nagel MN, Germany), and phosphorus concentrations were calculated by
interpolation from the standard calibration curve. The calibration curve
demonstrated strong linearity, with a correlation coefficient (R2) of 0.99.
Each measurement was duplicate, and assay precision was confirmed with relative
standard deviation (RSD) values below 5%. No matrix interference or blank signal
irregularities were observed. Outlier screening was conducted using a
All continuous data were tested for normality using the Shapiro–Wilk test and
for homogeneity of variance using Levene’s test. Differences between groups were
assessed using one-way ANOVA with Tukey’s Honestly Significant Difference (HSD)
post hoc test for pairwise comparisons. When assumptions were not met,
non-parametric alternatives were applied. Potential outliers were identified a
priori as values exceeding
Table 2 illustrates the comparison of dietary phosphorus content and intake
among the experimental groups. No significant differences were observed between
the S and O control groups regarding phosphorus content, daily food consumption,
and daily phosphorus intake, confirming comparable baseline conditions. Among the
intervention groups, the OL group showed a significantly higher dietary
phosphorus content compared with the O group (95% CI: 77.5 to 285; p
| Group | Parameter/unit | Mean |
Q1 | Q3 | IQR |
| S | Phosphorus | 1098 |
7.7 | 9.0 | 1.4 |
| O | (mg/kg diet d.w.) | 1098 |
8.0 | 9.3 | 1.4 |
| OB | 1185 |
8.0 | 8.3 | 0.4 | |
| OD | 1060 |
8.0 | 9.0 | 1.1 | |
| OT | 1198 |
7.7 | 8.0 | 0.3 | |
| OL | 1279 |
7.8 | 8.1 | 0.3 | |
| ODL | 1035 |
7.7 | 8.7 | 1.0 | |
| OTL | 1121 |
8.0 | 8.4 | 0.5 | |
| S | Food Intake | 16,754 |
16,007 | 16,939 | 933 |
| O | (mg/day) | 17,782 |
17,074 | 18,000 | 925 |
| OB | 17,265 |
17,092 | 17,383 | 291 | |
| OD | 17,982 |
17,338 | 18,684 | 1346 | |
| OT | 16,802 |
16,120 | 17,164 | 1045 | |
| OL | 17,538 |
16,516 | 18,034 | 1518 | |
| ODL | 16,931 |
16,208 | 17,038 | 830 | |
| OTL | 16,668 |
16,159 | 16,846 | 687 | |
| S | Phosphorus Intake | 18.4 |
17.6 | 18.6 | 1.0 |
| O | (mg/day) | 19.5 |
18.8 | 19.8 | 1.0 |
| OB | 20.5 |
20.3 | 20.6 | 0.4 | |
| OD | 19.1 |
18.4 | 19.8 | 1.4 | |
| OT | 20.1 |
19.3 | 20.6 | 1.3 | |
| OL | 22.4 |
21.1 | 23.1 | 1.9 | |
| ODL | 17.5 |
16.8 | 17.6 | 0.9 | |
| OTL | 18.7 |
18.1 | 18.9 | 0.8 |
S, Sham group receiving AIN-93M; O, Ovariectomized (OVX) group receiving
AIN-93M; OB, OVX group receiving AIN-93M with alendronate; OD, OVX group
receiving AIN-93M with daidzein; OT, OVX group receiving AIN-93M with tempeh; OL,
OVX group receiving AIN-93M with probiotics; ODL, OVX group receiving AIN-93M
with daidzein and probiotics; OTL, OVX group receiving AIN-93M with tempeh and
probiotics. Statistical analysis was conducted using ANOVA, followed by Tukey’s
post hoc test to determine significant differences between groups. Data are
expressed as mean
Table 3 summarizes the effects of dietary interventions on body mass after the six-week experimental period. Among the control groups, the O group exhibited a significantly higher body mass compared with the S group (95% CI: 8.05 to 103; p = 0.01). No significant differences were observed among the treated OVX groups; however, when compared with the S group, both the OD (95% CI: 4.12 to 95.9; p = 0.02) and OL (95% CI: 0.87 to 92.6; p = 0.04) groups displayed significantly higher body mass values.
| Group | Body mass (g) | |||
| Mean |
Q1 | Q3 | IQR | |
| S | 311 |
298 | 331 | 33.0 |
| O | 366 |
351 | 374 | 23.0 |
| OB | 356 |
329 | 376 | 46.5 |
| OD | 361 |
350 | 372 | 22.5 |
| OT | 354 |
335 | 366 | 30.8 |
| OL | 358 |
338 | 377 | 39.5 |
| ODL | 345 |
321 | 367 | 45.3 |
| OTL | 350 |
327 | 364 | 36.3 |
S, Sham rats receiving AIN-93M; O, Ovariectomized (OVX) rats receiving AIN-93M;
OB, OVX rats receiving AIN-93M with alendronate; OD, OVX rats receiving AIN-93M
with daidzein; OT, OVX rats receiving AIN-93M with tempeh; OL, OVX rats receiving
AIN-93M with probiotics; ODL, OVX rats receiving AIN-93M with daidzein and
probiotics; OTL, OVX rats receiving AIN-93M with tempeh and probiotics.
Statistical analysis was conducted using ANOVA, followed by Tukey’s post hoc test
to determine significant differences between groups. Data are expressed as
mean
Fig. 1 summarizes phosphorus levels across feces and tissue samples. No
significant differences in phosphorus levels were observed between the S and O
control groups in any of the analyzed tissues. Significant differences in
phosphorus levels were observed among different organs in the treated groups. In
femoral tissue, the OT group exhibited a significantly higher phosphorus level
compared with the O group (95% CI: 3.90 to 66.0; p = 0.01),
corresponding to an increase of roughly 25%. In the kidney, phosphorus levels of
the ODL group were elevated compared to the S group (95% CI: 157 to 1874;
p = 0.01), though differences compared to the O group did not reach
statistical significance. In the pancreas, phosphorus levels were significantly
higher in the OD (95% CI: 42.8 to 335; p
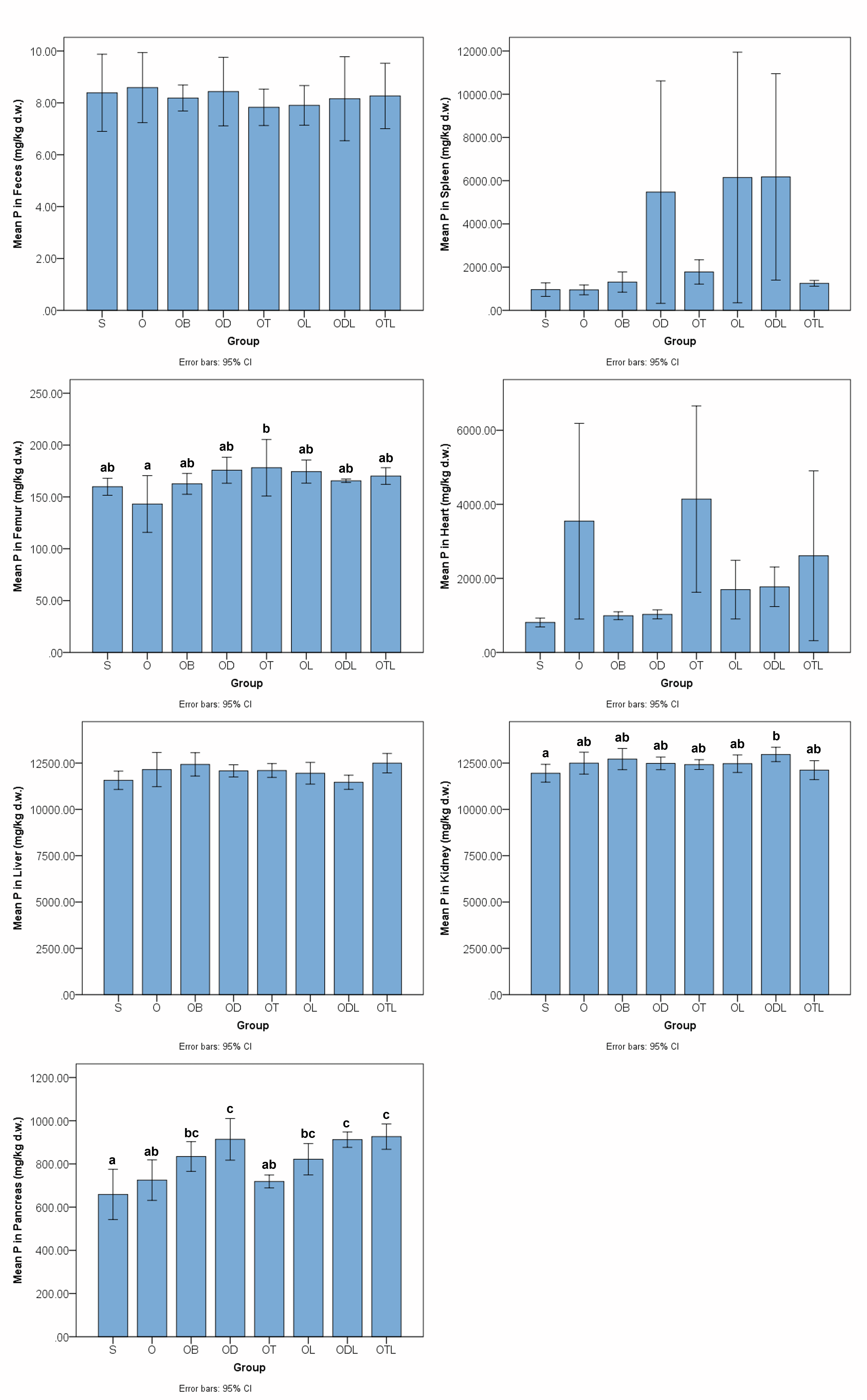 Fig. 1.
Fig. 1.
Phosphorus levels in feces and tissues after the six-week
intervention with various modified diets. Data are presented as bar plots
representing mean values
Table 4 presents Pearson’s correlation coefficients between feces and tissue phosphorus concentrations after six weeks of dietary intervention. No significant correlations were detected among most evaluated tissues (feces, spleen, femur, heart, liver, kidney, and pancreas). However, as illustrated in Fig. 2, a positive correlation was observed between pancreatic and femoral phosphorus levels, with a correlation coefficient of 0.33 and a significance level of 0.02. This finding indicates that higher phosphorus accumulation in the pancreas was associated with greater deposition in bone.
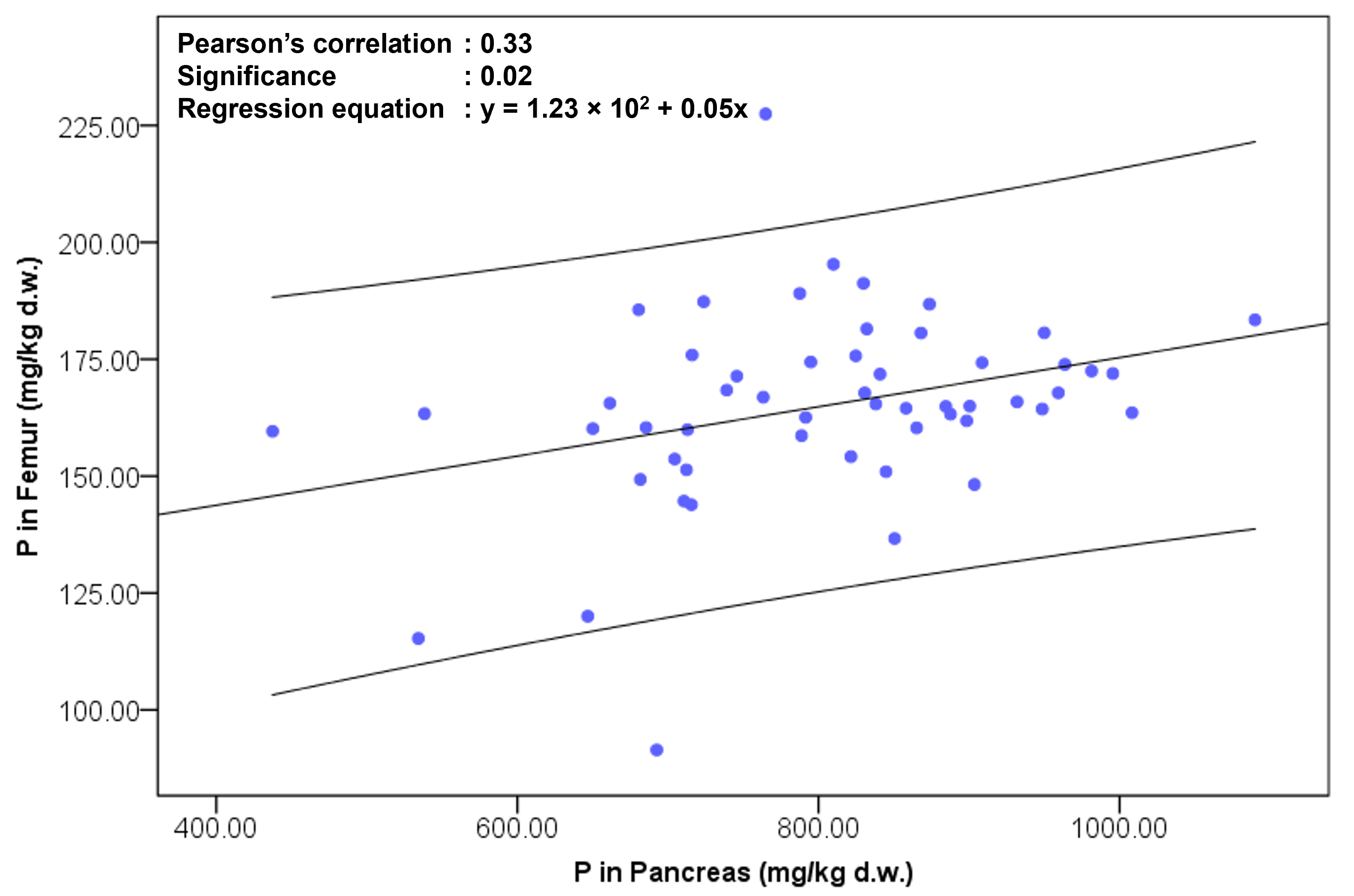 Fig. 2.
Fig. 2.
Pearson’s correlation between phosphorus (P) concentrations in
the femur and pancreas following a six‑week dietary intervention. Each data
point represents an individual sample (blue circles). The x‑axis shows P
concentration in the pancreas (mg/kg dry weight), and the y‑axis shows P
concentration in the femur (mg/kg dry weight). A linear regression line with 95%
confidence intervals is fitted to the data. The regression equation is y = 1.23
| Pair | Correlation coefficient | Significance |
| Feces–Spleen | –0.36 | 0.07 |
| Feces–Femur | –0.05 | 0.80 |
| Feces–Heart | 0.09 | 0.65 |
| Feces–Liver | –0.05 | 0.80 |
| Feces–Kidney | 0.26 | 0.17 |
| Feces–Pancreas | –0.26 | 0.18 |
| Spleen–Femur | 0.19 | 0.18 |
| Spleen–Heart | –0.12 | 0.40 |
| Spleen–Liver | –0.17 | 0.21 |
| Spleen–Kidney | 0.12 | 0.39 |
| Spleen–Pancreas | 0.20 | 0.14 |
| Femur–Heart | –0.26 | 0.06 |
| Femur–Liver | –0.21 | 0.12 |
| Femur–Kidney | 0.10 | 0.48 |
| Heart–Liver | 0.10 | 0.45 |
| Heart–Kidney | –0.03 | 0.85 |
| Heart–Pancreas | –0.20 | 0.14 |
| Liver–Kidney | –0.15 | 0.25 |
| Liver–Pancreas | 0.09 | 0.49 |
| Kidney–Pancreas | 0.18 | 0.16 |
This study was designed to investigate the effects of dietary interventions, specifically daidzein, tempeh, and L. acidophilus, on phosphorus metabolism in a postmenopausal rat model. Our findings indicate that the dietary interventions yielded differential effects on phosphorus levels among the examined tissues. Notably, the femur, kidney, and pancreas exhibited significant increases in phosphorus deposition following treatment. Furthermore, a significant positive correlation was observed between pancreatic and femoral phosphorus levels, suggesting a potential systemic link in mineral handling under dietary modulation. This study provides novel insights into the ability of phytoestrogen- and probiotic-based interventions to modulate systemic phosphorus metabolism in ovariectomized animals.
The primary finding of this study (Fig. 1) is that the femur, kidney, and pancreas were the most responsive tissues to dietary treatments with daidzein, tempeh, and L. acidophilus. These outcomes are consistent with previous research on calcium metabolism, where modified diets were shown to alter mineral distribution in serum and bone tissues. Specifically, previous studies demonstrated that dietary interventions such as tempeh and probiotics reduced serum calcium levels while enhancing femoral calcium deposition [6], suggesting a shift in mineral allocation from circulation to bone tissue. The current findings extend this concept to phosphorus, indicating that bioactive dietary components not only modulate calcium handling but also promote phosphorus deposition in skeletal and metabolically active organs.
The parallel responses of calcium and phosphorus may be partly explained by shared transport and regulatory mechanisms. Earlier investigations showed that dietary treatments altered the expression of calcium transporters, including TRPV5 (transient receptor potential vanilloid 5) and TRPV6 (transient receptor potential vanilloid 6), in the small intestine, with reduced expression in some groups and compensatory upregulation in others [7]. Although these transporters are specific to calcium, their modulation could indirectly influence phosphorus availability, given the interdependence of calcium–phosphorus absorption and homeostasis. Furthermore, the observed increase in feces calcium excretion in some treatment groups corresponds with the present lack of significant feces–tissue phosphorus correlations, underscoring that mineral redistribution in response to bioactive compounds may be tissue-specific rather than systemic.
Building on these observations, these results demonstrate that daidzein, tempeh, and L. acidophilus act in concert to influence calcium and phosphorus metabolism, thereby contributing to mineral equilibrium in the postmenopausal state. The marked increases in phosphorus deposition within the femur, kidney, and pancreas point to tissue-specific responsiveness, suggesting that both skeletal and metabolic compartments participate in dietary regulation of mineral balance. This coordinated response highlights a broader systemic interplay, in which dietary bioactives modulate not only bone mineralization but also extra-skeletal phosphorus handling. These findings extend the understanding of postmenopausal mineral metabolism and underscore the translational potential of dietary strategies for improving mineral regulation and skeletal health.
In the femoral bone, adequate phosphorus availability, in combination with calcium, is critical for hydroxyapatite formation and bone mineralization [28], and the observed improvements may reflect enhanced bone matrix synthesis under conditions of estrogen deficiency [29]. In the kidney, estrogen deficiency can impair phosphate excretion and renal handling, mediated via changes in FGF23 and PTH signaling [30]. In the pancreas, phosphate is essential for ATP synthesis, enzyme activation, and insulin secretion, suggesting that these dietary interventions could impact metabolic regulation in estrogen-deficient states [31].
These findings demonstrate that dietary interventions with daidzein, tempeh, and L. acidophilus distinctly influenced phosphorus metabolism in a postmenopausal rat model. Administration of pure daidzein increased pancreatic phosphorus content, which may be attributed to phytoestrogenic modulation of mineral transporters [32] and enhanced intestinal absorption [33]. Importantly, daidzein was provided in its aglycone form, which is more bioavailable than the glycoside-conjugated isoflavones naturally present in tempeh [34]. Variations in chemical form may contribute to differences in how phosphorus is metabolized and distributed across organs.
Diets containing tempeh significantly associated with femoral phosphorus content compared to OVX controls. This effect may result from the combined action of bioactive isoflavones, fermentation-derived peptides, and improved mineral bioaccessibility [35]. Fermentation processes are known to enhance the solubility and absorption of calcium and phosphorus [35], which may explain the observed increases in bone mineral deposition.
Differences between daidzein alone and tempeh highlight the importance of the food matrix. Glycoside hydrolysis by intestinal microbiota in tempeh diets releases aglycone forms [8], while fermentation-derived peptides may further facilitate mineral uptake [36]. Although both interventions increase phosphorus availability, the matrix of tempeh appears to favor skeletal deposition, whereas pure daidzein preferentially affects pancreatic phosphorus. The divergent tissue patterns—greater femoral phosphorus accumulation following tempeh intake versus preferential pancreatic phosphorus increases with isolated daidzein—are plausibly explained by differences in food matrix, isoflavone speciation, and host–microbial interactions. Fermentation transforms soy into a complex matrix that contains bioactive peptides, converted aglycones, and reduced phytate levels [8]; these changes enhance overall mineral bioaccessibility and may promote intestinal uptake of phosphate in a form that is preferentially deposited in bone. In addition, tempeh supplies macronutrients and peptide factors that may favor skeletal anabolism and mineral incorporation, thereby biasing newly absorbed phosphate toward the femur. By contrast, the pure daidzein provided here in aglycone form is rapidly absorbed and exerts direct phytoestrogenic and metabolic signaling effects that may modulate pancreatic cellular processes [37]—potentially altering local phosphate transport or retention. Finally, probiotic co-administration may further modify these trajectories through microbial phytase activity and altered metabolite profiles [38], thereby shifting both the amount and the kinetics of phosphate available for organ uptake. Taken together, these mechanisms provide a coherent explanation for why a whole-food, fermented matrix tends to favor skeletal phosphate deposition, whereas an isolated, rapidly absorbed isoflavone may preferentially influence metabolically active organs such as the pancreas; targeted mechanistic studies are warranted to test these hypotheses.
Rats receiving L. acidophilus exhibited significantly higher dietary phosphorus content and intake. This effect may be attributed to microbial activity facilitating the release of phosphorus from phytates and other mineral-bound complexes within the diet, thereby enhancing intestinal absorption. Recent advances in probiotic research emphasize the importance of targeted delivery and encapsulation techniques in enhancing probiotic survival and intestinal efficacy, thereby improving host nutrient absorption and metabolic regulation [39]. Mechanistically, probiotics can modulate gut microbiota, release enzymes such as phytases, and enhance intestinal absorption of phosphorus and calcium [40, 41]. When combined with isoflavone-containing diets, probiotics may also influence bile acid metabolism [42], improve mineral solubility, and support mineral deposition [43]. These interactions underscore the potential of dietary bioactives and microbial modulation to influence systemic phosphorus homeostasis in estrogen-deficient models.
Although these interventions yielded promising outcomes individually, no evident synergistic effects were observed when daidzein, tempeh, and probiotics were combined. This lack of synergy may be attributed to overlapping mechanisms of action, saturation of phosphorus transport and storage pathways, or limited bioavailability of isoflavones and microbial metabolites under the combined treatment conditions.
Second, OVX rats exhibited a significant association with body mass compared to sham-operated controls (Table 3), consistent with the well-established effects of estrogen deficiency on adiposity and metabolic regulation [44]. Mechanistically, estrogen loss promotes hyperphagia-independent weight gain through reduced energy expenditure [45], altered lipid metabolism [46], and enhanced adipocyte differentiation, particularly in visceral depots [47]. Additionally, estrogen deficiency shifts the balance of anabolic and catabolic signaling in skeletal muscle and liver, favoring lipid accumulation and altered glucose utilization, which collectively contribute to increased body mass [48].
Despite this weight gain, no significant differences were observed in daily dietary phosphorus intake between the OVX and sham groups (Table 2). This suggests that the observed increase in body mass was not driven by differences in phosphorus consumption. Mechanistically, phosphate homeostasis is tightly regulated by endocrine and renal mechanisms, including FGF23, PTH, and vitamin D-mediated intestinal absorption and renal reabsorption [49]. These regulatory pathways maintain stable systemic and tissue phosphorus concentrations even under conditions of increased bone resorption associated with estrogen deficiency [50].
Alendronate treatment was included as a pharmacological reference to provide context for evaluating the efficacy of diet-based strategies on phosphorus metabolism, given its well-established role in inhibiting osteoclast-mediated bone resorption and clinical relevance in postmenopausal osteoporosis management [51]. In the present study, however, no significant differences in phosphorus concentrations across tissues, body mass, or other measured parameters were observed between alendronate-treated, ovariectomized, and sham-operated groups. This finding contrasts with reports describing alendronate-induced improvements in bone mineral content [52]. Several factors may account for the observed outcome. One important consideration is the applied dose of alendronate (3 mg/kg/week), which may not have been sufficient to elicit measurable alterations in phosphorus distribution. A previous report has demonstrated variability in treatment responses depending on both dosage and frequency of administration [52]. For example, the report has described dosing regimens ranging from 0.1 mg/kg administered repeatedly over an extended period to 7 mg/kg given less frequently, both of which were shown to enhance systemic alendronate delivery and improve osseointegration. However, despite these findings, the optimal dose and frequency required to achieve favorable systemic and tissue-specific effects remain unresolved [52]. In addition, the postmenopausal rat model may exhibit different responsiveness compared to human physiology [53]. Moreover, phosphorus homeostasis alone may not fully capture the pharmacological effects of alendronate treatment, since these compounds primarily act by suppressing osteoclast-mediated bone resorption and indirectly supporting bone formation, rather than exerting direct regulatory control over systemic phosphorus levels [54]. In contrast, dietary interventions with daidzein, tempeh, and probiotics resulted in measurable effects on phosphorus distribution, suggesting that bioactive compounds can modulate mineral metabolism through gut-associated mechanisms distinct from the antiresorptive pathway targeted by alendronate.
Another noteworthy finding in this study was the positive correlation observed between pancreatic and femoral phosphorus concentrations, suggesting a potential systemic coordination between bone mineralization and pancreatic phosphorus metabolism. This association is particularly intriguing, as links between skeletal mineral dynamics and pancreatic function have rarely been reported in the literature. This relationship may be mediated by endocrine factors such as FGF23, which regulates phosphate homeostasis through its actions on renal excretion and vitamin D metabolism [55]. Given that insulin and IGF-1 are known to suppress FGF23 production via the PI3K/Akt/FOXO1 pathway, pancreatic function could indirectly influence skeletal phosphate regulation. The interplay among insulin, IGF-1, and FGF23 therefore provides a plausible mechanistic link connecting pancreatic phosphorus metabolism with bone mineral dynamics [55]. While the underlying mechanisms remain to be determined, this association generates hypotheses regarding possible regulatory interactions between bone mineralization and pancreatic function. Endocrine factors such as FGF23, which regulate phosphate homeostasis through renal excretion and vitamin D metabolism, as well as insulin and IGF-1, may be involved in this axis. However, these mechanisms were not directly evaluated in the present study, and further investigations are warranted. This observation highlights a novel, underexplored relationship that could inform future studies aimed at understanding systemic mineral regulation and the impact of dietary bioactive on bone and metabolic health.
Recent evidence has highlighted the complex endocrine network regulating phosphate balance, primarily mediated through the FGF23–PTH–vitamin D axis. FGF23, secreted mainly by osteocytes, acts on the kidneys to promote phosphate excretion and suppress 1,25(OH)2D synthesis, thereby maintaining systemic phosphate homeostasis [56]. Crosstalk between phosphate sensing, iron metabolism, and FGF23 regulation has also been described, revealing novel pathways that link mineral metabolism to systemic energy status [57]. Moreover, the dynamic interaction between FGF23, PTH, and vitamin D forms a tightly regulated bone–kidney–parathyroid feedback loop essential for mineral balance and skeletal integrity [58]. These insights provide a mechanistic foundation for interpreting the phosphorus-related changes observed in our study and underscore the importance of endocrine regulators in bone–phosphate metabolism.
Moreover, the translational relevance of these findings lies in their potential implications for postmenopausal women, in whom disturbances in mineral metabolism contribute to the risk of osteoporosis and related comorbidities. Dietary sources rich in isoflavones, such as tempeh, together with probiotics, may offer a sustainable nutritional strategy to support phosphorus homeostasis and bone integrity through gut-mediated and endocrine-modulating pathways. At the same time, it is essential to recognize the importance of maintaining an appropriate calcium-to-phosphorus ratio in dietary interventions aimed at supporting bone health. Together, these findings highlight the promise of food-based approaches as complementary to conventional therapies, while also emphasizing the necessity of carefully balancing benefits with potential systemic risks.
A major strength of the present study lies in its integrative design, which compared the effects of an isolated phytoestrogen, a fermented soy-based food, and a probiotic strain, thereby enabling the distinction between compound-specific, matrix-dependent, and microbiota-mediated effects on phosphorus metabolism. The inclusion of multiple tissues, rather than limiting the analysis to bone, provided a broader perspective on systemic phosphorus handling, and the correlation analysis offered novel insights into inter-organ relationships, such as the pancreas–bone axis.
Nevertheless, several limitations should be acknowledged. The dose-response relationships for daidzein, tempeh, and L. acidophilus were not systematically evaluated; therefore, the optimal concentrations required to maximize phosphorus modulation without affecting overall mineral balance remain undetermined. The alendronate group served as a pharmacological reference rather than confirmatory control, as exposure verification was not performed. Therefore, its outcomes should be interpreted descriptively, recognizing potential biological variability and differences in pharmacodynamic response. Furthermore, the present study focused on phosphorus distribution at the tissue level and did not include systemic markers such as FGF23, PTH, or vitamin D metabolites, which could clarify endocrine mechanisms underlying phosphorus regulation. Future studies should integrate these biomarkers to establish causal relationships between dietary interventions and systemic phosphate homeostasis. As dietary intake was measured per cage rather than per individual, statistical analyses were based on individual animal outcomes. This introduces a potential risk of pseudo-replication, which may slightly inflate Type I error rates. Consequently, all correlation analyses should be interpreted as exploratory. Moreover, although dietary phosphorus and Ca:P ratios were analyzed and found to be comparable among groups, minor differences cannot be fully excluded and may have contributed to observed tissue phosphorus variability. This factor should be considered when interpreting between-group effects. Addressing these aspects in future work, ideally through longer-term interventions and the inclusion of human trials, will be crucial to fully establish the translational relevance of these findings.
Dietary interventions with daidzein, tempeh, and L. acidophilus significantly increased phosphorus concentrations in the femur, kidney, and pancreas of a postmenopausal rat model. These outcomes extend previous evidence of calcium redistribution under similar interventions, underscoring the interdependence of calcium and phosphorus metabolism in the regulation of mineral balance. The parallel increases in tissue phosphorus and femoral calcium suggest that dietary bioactives promote coordinated deposition across skeletal and metabolic organs. Although no consistent synergistic effects among daidzein, tempeh, and probiotics were identified, the positive correlation between pancreatic and femoral phosphorus points to systemic mechanisms that link bone mineralization with extra-skeletal phosphorus handling. Collectively, these findings provide novel evidence that targeted dietary bioactives may support phosphorus homeostasis and bone mineralization, offering translational potential for nutritional strategies aimed at improving mineral metabolism in postmenopausal women.
Raw per-animal data on phosphorus intake, tissue phosphorus concentrations, and body weight, as well as the analysis code, are available in the REPOd data repository (https://doi.org/10.18150/QHJUSA).
IAH and JS conceptualized and designed the research study. IAH conducted the investigation, curated the data, performed the formal analysis, acquired funding, administered the project, provided resources, and prepared the visualizations. IAH, KS, PK, and JS developed the methodology. JS supervised the study. IAH and JS validated the results. IAH wrote the original draft of the manuscript. IAH and JS reviewed and edited the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
The animal study protocol was approved by the Local Ethical Committee (Lokalna Komisja Etyczna) in Poznan, Poland (No. registration: 21/2021, Issued date: 21 May 2021). Experiments followed EU Directive 2010/63, NIH guidelines for laboratory animal care, and the ARRIVE reporting principles. The completed checklist is provided in a supplementary file. Animal handling adhered to institutional standards at the Poznań University of Life Sciences, ensuring full compliance with national and international regulations.
We would like to express our gratitude to Professor Jerzy Nawracała for providing the soybean used in this study.
This study was funded by the Polish National Science Centre/Narodowe Centrum Nauki (Grant no.: 2021/41/N/NZ9/00838; Grant holder: Iskandar Azmy Harahap).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/IJVNR46717.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.