

 , Zhuo Zhang 1,2, Xin Xiao 3, Xu Leng 1,2, Junru He 1,2, Shaowu Xiao 1,2, Mengya Zeng 1,*
, Zhuo Zhang 1,2, Xin Xiao 3, Xu Leng 1,2, Junru He 1,2, Shaowu Xiao 1,2, Mengya Zeng 1,* , Yuewu Chen 1,*
, Yuewu Chen 1,*1 Department of Cardiology, Second Affiliated Hospital of Hainan Medical University, 570311 Haikou, Hainan, China
2 Second Clinical Medical College, Hainan Medical University, 571199 Haikou, Hainan, China
3 Medical College, Jishou University, 416000 Jishou, Hunan, China
Abstract
Cardiovascular diseases (CVD) and atherosclerosis (AS) are major global health issues. Bisphenol A (BPA), a common endocrine-disrupting chemical, has been closely linked to the development of AS with long-term exposure. This study aims to explore the molecular mechanisms underlying BPA-induced AS using network toxicology, molecular docking, and machine learning approaches.
First, Absorption, Distribution, Metabolism, Excretion, and Toxicity (ADMET) analysis of BPA was conducted to predict its absorption, metabolism, and toxicity characteristics in the body. Subsequently, potential targets of BPA related to AS were predicted using public databases, followed by Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis to identify relevant signaling pathways. Core genes were selected using CytoNCA plugin, Least Absolute Shrinkage and Selection Operator (LASSO) regression, and Support Vector Machine Recursive Feature Elimination (SVM-RFE) machine learning algorithms. Finally, validation was performed using receiver operating characteristic (ROC) curves and molecular docking.
ADMET analysis showed that BPA may promote vascular remodeling and inflammatory responses under long-term low-dose exposure, exacerbating the development of AS. Network toxicology analysis predicted 372 intersecting genes related to BPA and AS. KEGG analysis indicated that BPA might contribute to AS progression through signaling pathways such as Phosphatidylinositol 3‑kinase–protein kinase B (PI3K‑Akt)and Mitogen‑Activated Protein Kinase (MAPK). Protein interaction network analysis identified 61 core genes, and machine learning approaches using LASSO and SVM methods identified 11 and 16 core genes, respectively, with TNF being the key core gene. ROC curve analysis demonstrated a strong diagnostic potential for TNF in AS, with an AUC value approaching 1.0, suggesting that TNF is a potential biomarker for AS. Molecular docking results showed a binding energy of –7.3 kcal/mol between BPA and TNF-α.
This study employed multiple technical approaches to identify TNF as a key core gene in BPA-induced AS and validated its diagnostic potential as a biomarker. BPA may promote AS development by binding to TNF-α and its signaling pathways, offering new therapeutic insights for TNF-targeted intervention strategies.
Keywords
- bisphenol A
- atherosclerosis
- tumor necrosis factor-alpha
- molecular docking simulation
- machine learning
Cardiovascular diseases (CVD) and atherosclerosis (AS) are major global health challenges, placing a significant burden on global public health [1, 2]. CVD is the leading cause of death worldwide, responsible for nearly 20 million deaths each year, and its prevalence is expected to increase by 90% by 2050 [3]. AS, as the primary pathological basis of CVD, is mainly caused by the accumulation of lipids, cholesterol, and other substances in the vascular walls, forming atherosclerotic plaques. This leads to the narrowing or blockage of blood vessels, severely affecting blood circulation and potentially causing serious complications such as myocardial infarction and stroke [4, 5, 6]. In recent years, environmental pollutants, particularly endocrine disruptors, have played an increasingly important role in the occurrence and development of AS [7, 8].
Bisphenol A (BPA), a common endocrine disruptor, is widely present in plastic products, food packaging materials, electronic devices, and everyday items. Its widespread distribution and persistence in the environment have made it a key focus in the study of the impact of environmental pollution on human health [9, 10]. Studies have shown that long-term exposure to BPA is closely associated with the occurrence of a range of metabolic diseases, including AS [11, 12]. BPA may play a significant role in the development of AS by affecting the endocrine system, inducing oxidative stress, and promoting inflammatory responses [13, 14, 15]. However, the specific mechanisms by which BPA induces AS remain unclear and require further investigation [16].
With the development of bioinformatics and molecular biology, network toxicology has emerged as a new research approach that can reveal the potential effects of environmental toxins on organisms by constructing networks of interactions between biomolecules [17, 18]. Combined with molecular docking techniques, the interactions between BPA and target proteins can be explored in depth, further elucidating its molecular mechanisms [19]. At the same time, the application of machine learning methods in biological data analysis is becoming increasingly widespread, helping to identify potential biomarkers and predict the impact of BPA on the occurrence of AS [20, 21]. This study aims to deeply explore the molecular mechanisms of BPA-induced AS based on network toxicology, molecular docking, and machine learning methods. It is expected to provide new theoretical insights into the molecular mechanisms of BPA-induced AS and offer new strategies for the prevention and treatment of related diseases.
We obtained the SMILES structure of BPA from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/) and performed ADMET (Absorption, Distribution, Metabolism, Excretion, and Toxicity) analysis using ADMETlab 2.0 (https://admetmesh.scbdd.com/) [22]. The platform provides comprehensive prediction functions, allowing the evaluation of BPA’s absorption in the body, hepatic metabolism, P-glycoprotein efflux pump substrate potential, inhibition of cytochrome P450 (CYP) enzymes, and other toxicity-related parameters [23].
We obtained the 2D structure and SMILES code of BPA from the PubChem database,
and performed target prediction using default parameters in PharmMapper
(https://www.lilab-ecust.cn/pharmmapper/index.html), SwissTargetPrediction
(https://www.swisstargetprediction.ch/), and CHEMBL
databases (https://www.ebi.ac.uk/chembl/), selecting “Homo sapiens” as the
species for screening [24]. By merging and removing duplicates, a list of
potential targets was obtained. Additionally, relevant targets were screened
using the keyword “AS” in the GeneCards (https://www.genecards.org/) and OMIM
(https://www.omim.org/) databases, and targets with a relevance score greater
than 1 were selected from GeneCards [25]. All potential targets were standardized
using the UniProt database. Next, Kyoto Encyclopedia of Genes and Genomes (KEGG)
enrichment analysis was performed through the Database for Annotation,
Visualization, and Integrated Discovery (DAVID) database
(https://davidbioinformatics.nih.gov/), with a screening criterion of p
We imported the intersecting targets into the STRING database, selected “Homo sapiens” as the species, and set the minimum interaction confidence to 0.900. After exporting the results as a TSV file, the data was imported into Cytoscape v3.9.1, where free nodes were excluded, and the CytoNCA (v2.1.6) plugin was used for filtering [27]. During the filtering process, six centrality metrics were calculated: Degree Centrality (DC), Betweenness Centrality (BC), Closeness Centrality (CC), Eigenvector Centrality (EC), Local Average Connectivity (LAC), and Network Centrality (NC). Nodes with values above the median in all six metrics were retained [28].
We obtained the GSE100927 dataset from the Gene Expression Omnibus (GEO)
database (69 diseased arterial tissues and 35 control arterial tissues, platform:
GPL17077). This dataset, derived from human arterial tissues, provides good
biological comparability. The diseased arterial tissues were defined as the
experimental group, and the normal control arterial tissues as the control group.
Batch effect correction and dataset integration were performed using R (v4.5.2),
and differential expression analysis was conducted using the limma package
(v3.66). Genes with
We employed two complementary machine learning algorithms: Least Absolute Shrinkage and Selection Operator (LASSO) regression and Support Vector Machine Recursive Feature Elimination (SVM-RFE) to identify core genes [30, 31]. LASSO regression is a regression analysis method used for variable selection and regularization, which helps reduce overfitting and improve prediction accuracy [32]. SVM-RFE, based on Support Vector Machine, recursively eliminates less important features and progressively identifies the most contributing features for the classification task [31]. Both methods employed 10-fold cross-validation for model training and parameter optimisation to ensure the model’s generalisation capability and stability.
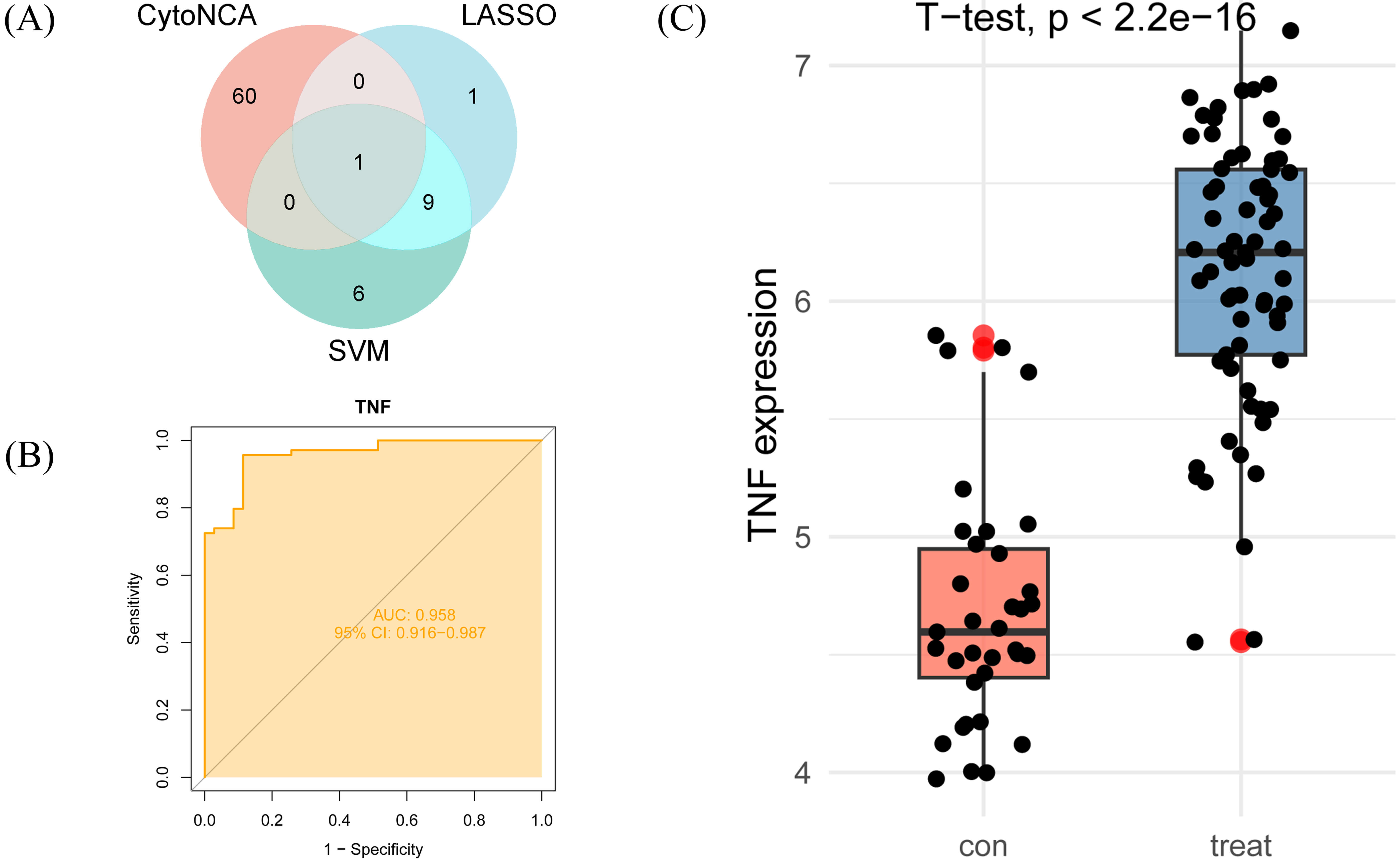
The key core genes were identified through the intersection of selections obtained via CytoNCA, LASSO regression, and SVM-RFE. We visualised these key core genes based on expression data from the control and AS groups within the GSE100927 dataset, and constructed receiver operating characteristic (ROC) curves to further evaluate their diagnostic potential [33]. Finally, calculate the area under the curve (AUC) to assess the diagnostic accuracy of the ROC curve [34].
We performed molecular docking validation on key core gene proteins, downloading the 3D structure of BPA from the PubChem database and obtaining the receptor’s structure file from the PDB database. A suitable docking box was customised according to the protein size, and docking was conducted using AutoDock Vina (v1.5.7) [35]. The lower the binding energy, the more stable the ligand-receptor complex. We visualised the protein-ligand conformation with the lowest binding energy using PyMOL 3.0 and LigPlot v.2.3.1 [36, 37].
Data were analyzed and visualized using R software (v4.5.2). Comparisons between
two groups with normal distributions were performed using the independent samples
t-test; comparisons between groups with non-normal distributions were
performed using the Wilcoxon signed-rank test. All tests were two-tailed, with
p
As an endocrine disruptor, BPA exhibits significant biotoxicity and metabolic
characteristics. ADMET analysis indicates (Supplementary Fig. 1) that
BPA possesses a Caco-2 permeability coefficient of –4.742, suggesting poor
intestinal absorption capacity. Its low Human Intestinal Absorption (HIA) value
further implies reduced oral absorption efficiency. The Madin-Darby Canine Kidney
(MDCK) permeability coefficient of 2
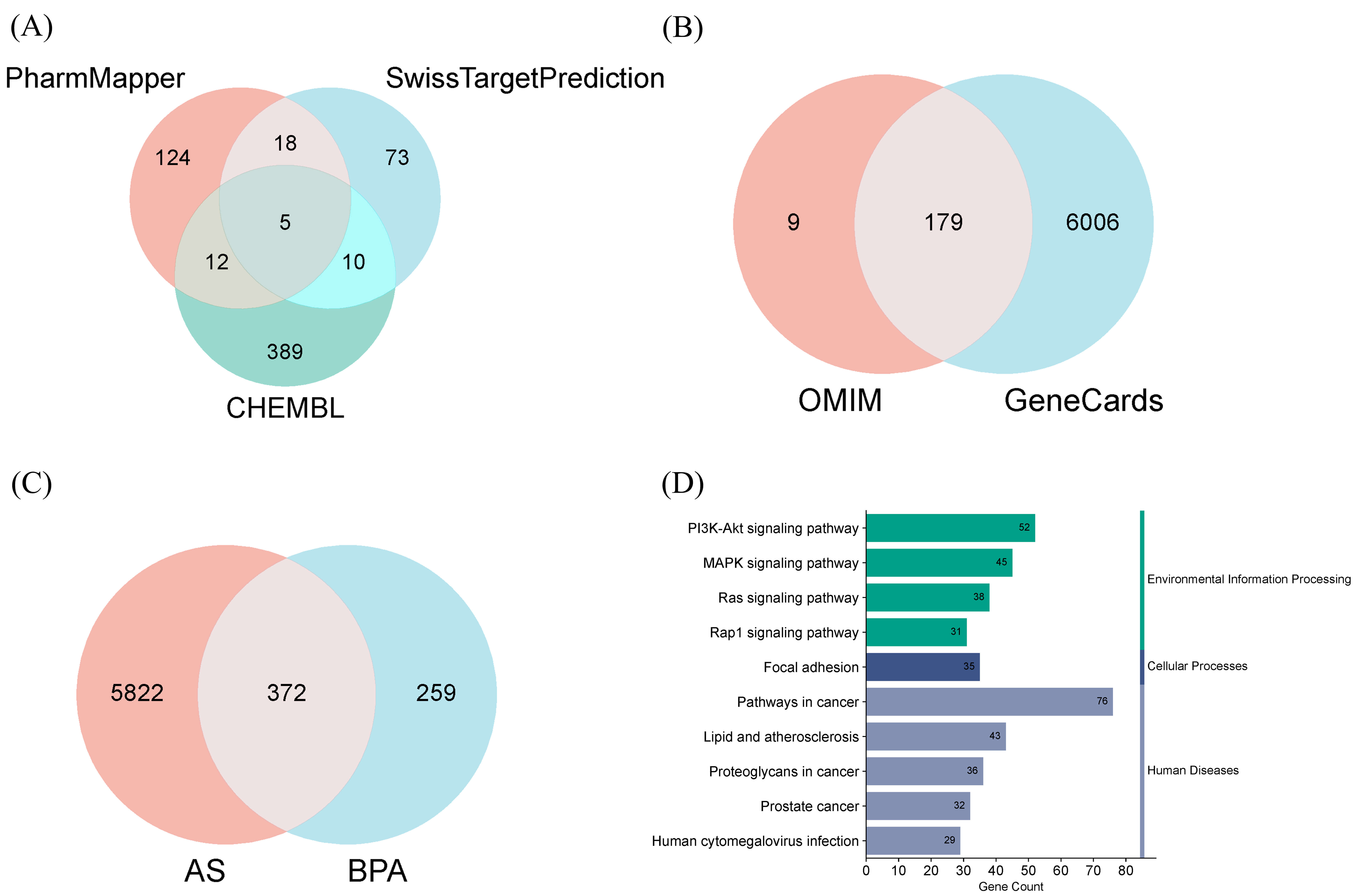
As shown in Fig. 1A, we predicted 159 potential targets using the PharmMapper
database, 106 using the SwissTargetPrediction database, and 416 using the CHEMBL
database (Supplementary Table 1). As depicted in Fig. 1B, we
additionally obtained 6185 targets from the GeneCards database and collected 188
targets from the OMIM database (Supplementary Table 2). An intersection
analysis of BPA-associated targets and AS-associated targets (Fig. 1C) revealed
372 overlapping genes. Subsequently, these 372 genes were imported into the DAVID
database for KEGG enrichment analysis, yielding 157 statistically significant
pathways (p
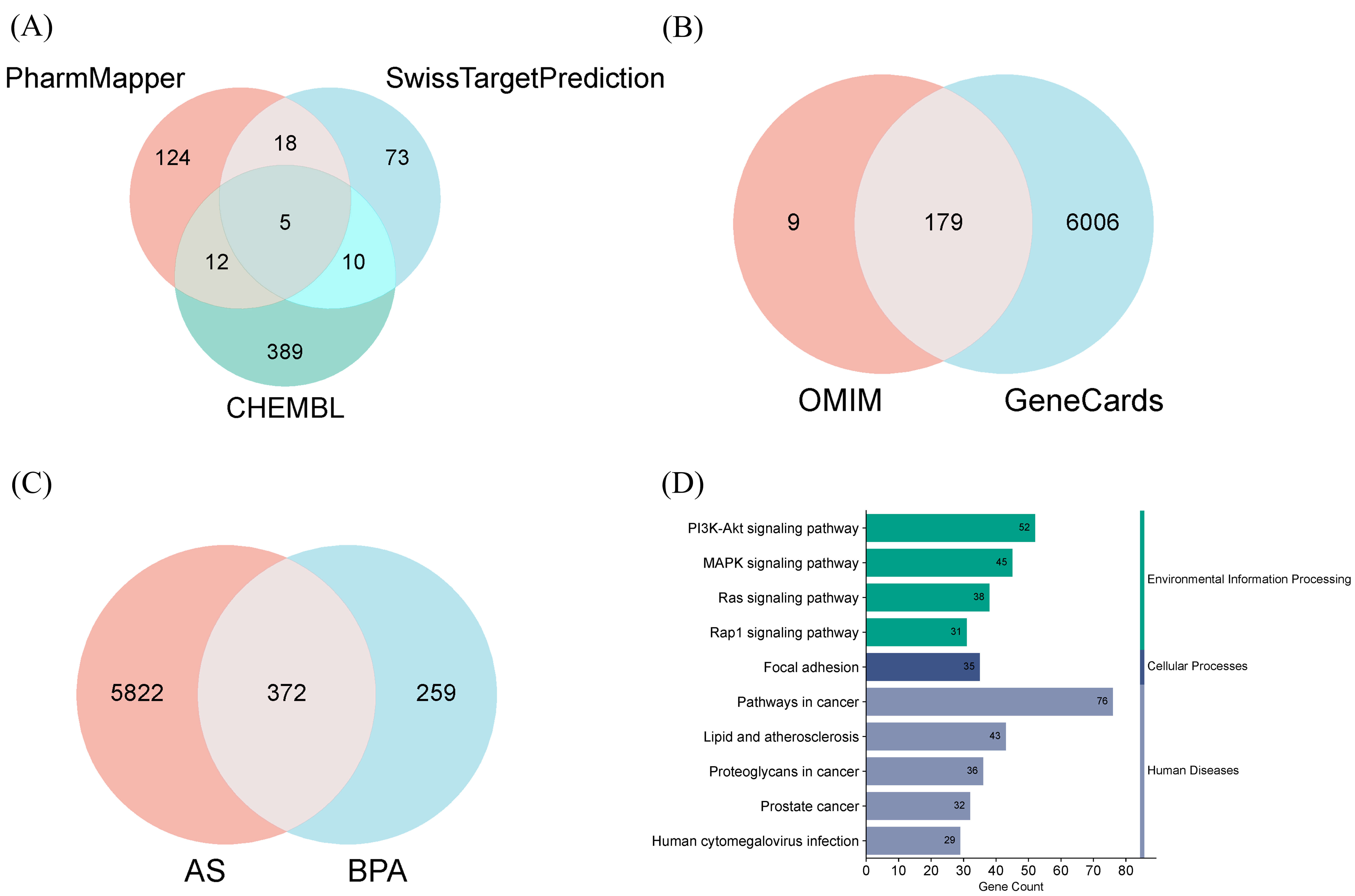 Fig. 1.
Fig. 1.
Identification of BPA-related and AS-related targets and KEGG enrichment analysis of overlapping genes. (A) Venn diagram of BPA targets. (B) Venn diagram of AS targets. (C) Venn diagram showing the intersection of BPA and AS targets. (D) Histogram of KEGG pathway classifications. BPA, Bisphenol A; AS, atherosclerosis; KEGG, Kyoto Encyclopedia of Genes and Genomes; OMIM, Online Mendelian Inheritance in Man.
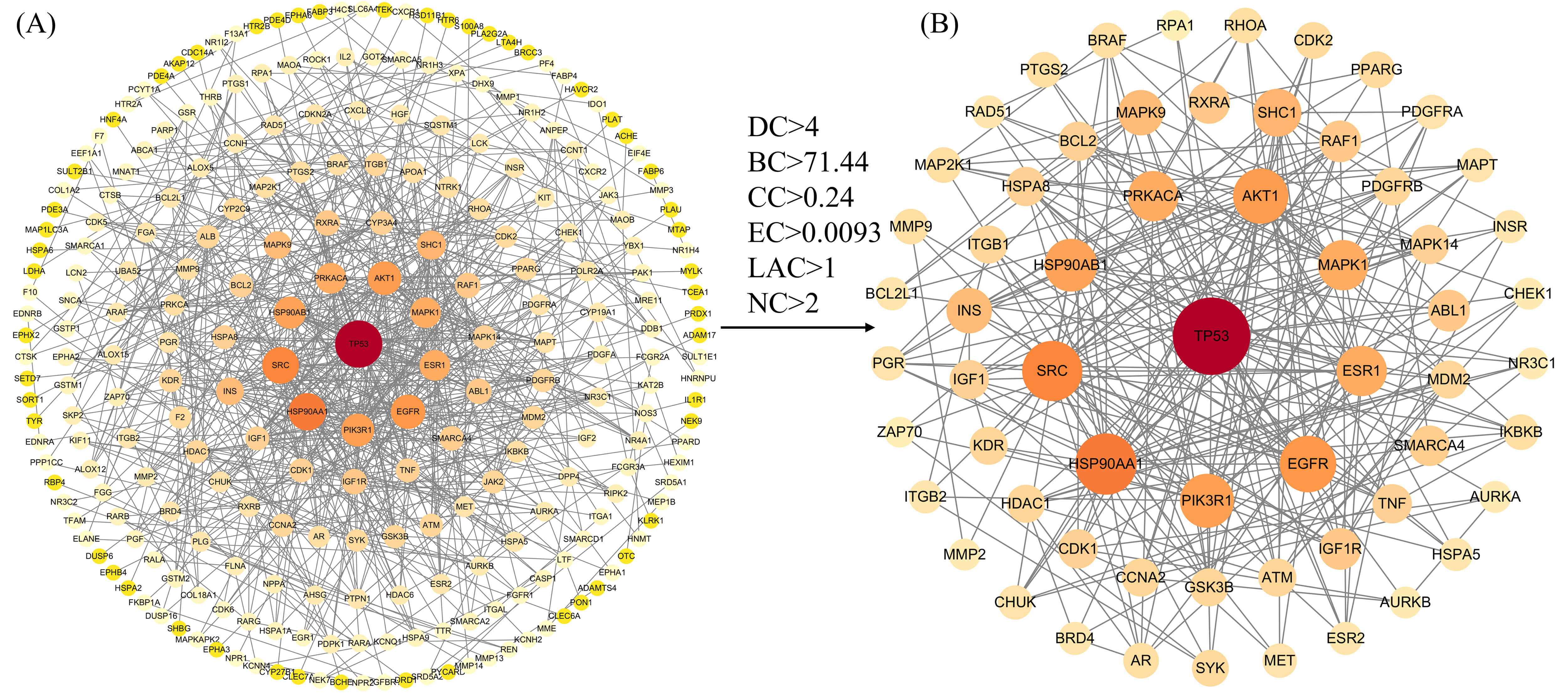
We constructed a protein interaction network comprising 372 intersecting genes using the STRING database, visualised via Cytoscape 3.10.3 software (Fig. 2A). This network contained 255 nodes and 691 edges, with an average node degree of 5.41; node size and colour varied according to degree values (Supplementary Table 4). Subsequently, median filtering was applied to the network using the CytoNCA plugin, yielding a subnetwork comprising 61 nodes and 383.5 edges (Fig. 2B). This subnetwork exhibits markedly enhanced “functional density”, demonstrating elevated values across DC, BC, CC, EC, LAC, and NC metrics, revealing characteristics such as hubs, bridges, and centres of influence (Supplementary Table 5).
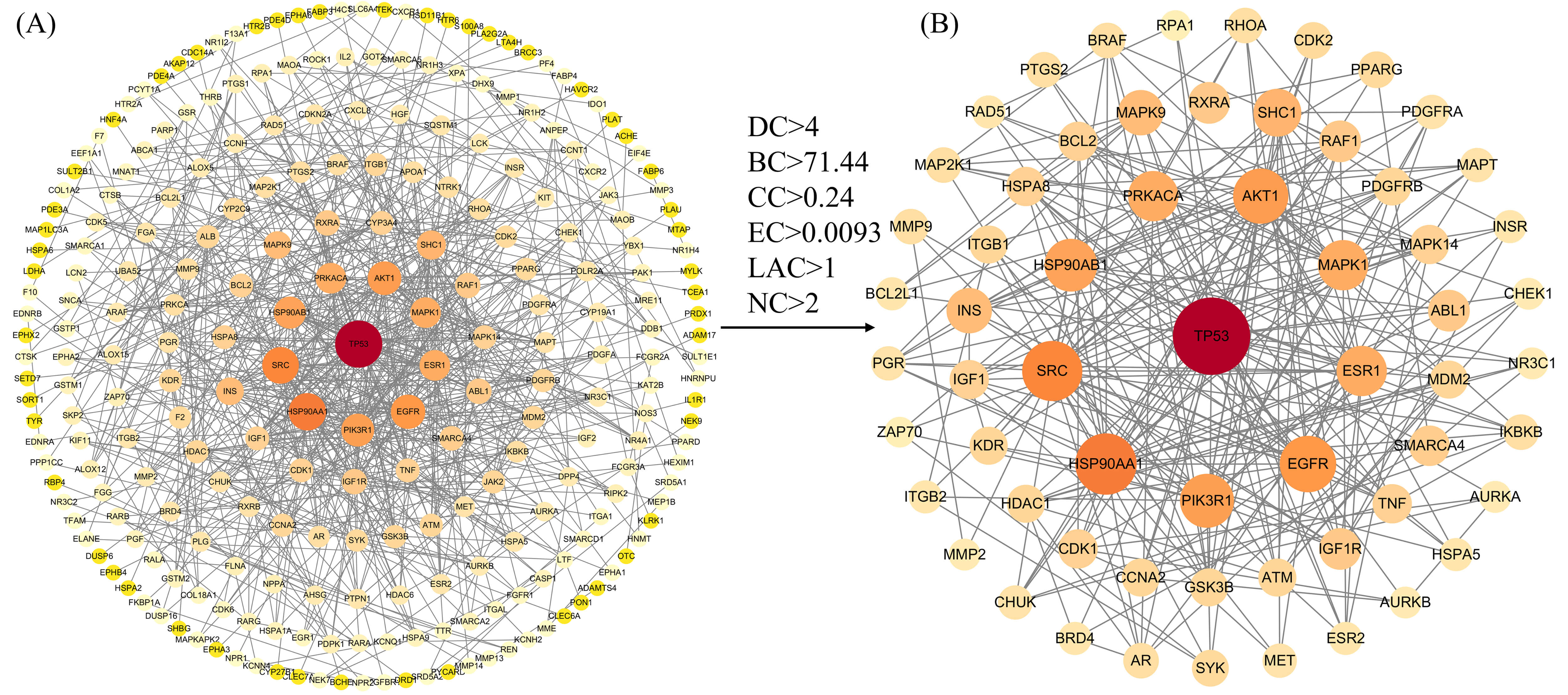 Fig. 2.
Fig. 2.
Protein-protein interaction network diagram. (A) The protein–protein interaction network before screening. (B) The core subnetwork obtained after median-based screening. The number of edges represents the sum of weighted edges. DC, Degree Centrality; BC, Betweenness Centrality; CC, Closeness Centrality; EC, Eigenvector Centrality; LAC, Local Average Connectivity; NC, Network Centrality.
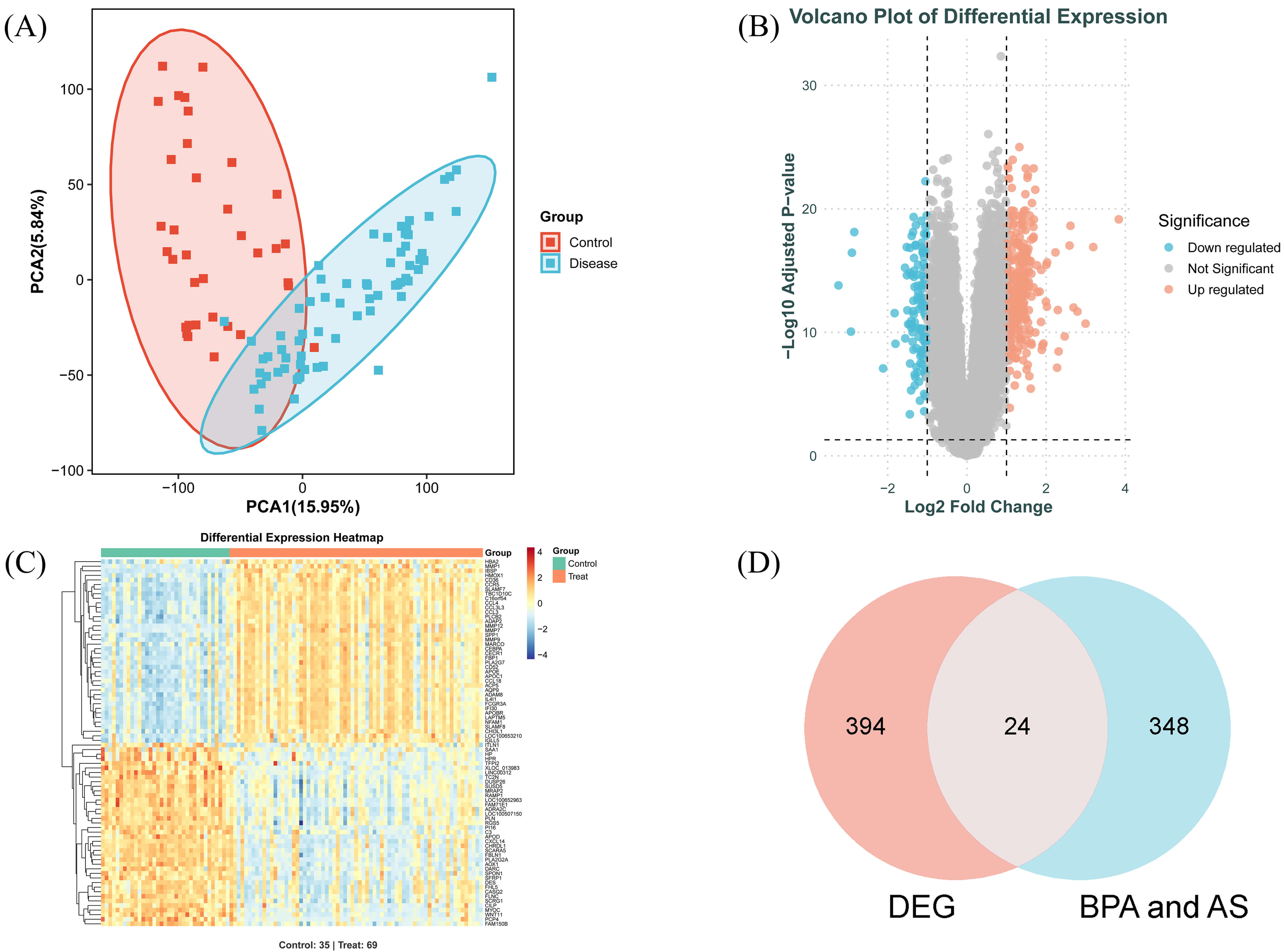
Following preprocessing and batch effect correction of the GSE100927 transcriptome dataset, a total of 418 differentially expressed genes (DEGs) were identified (Fig. 3B,C). Principal component analysis (PCA) (Fig. 3A) revealed significant differences between the normal and diseased groups. Intersection analysis of the 418 DEGs with the previously identified 372 genes yielded 24 common target genes, which were further utilised for machine learning screening (Fig. 3D).
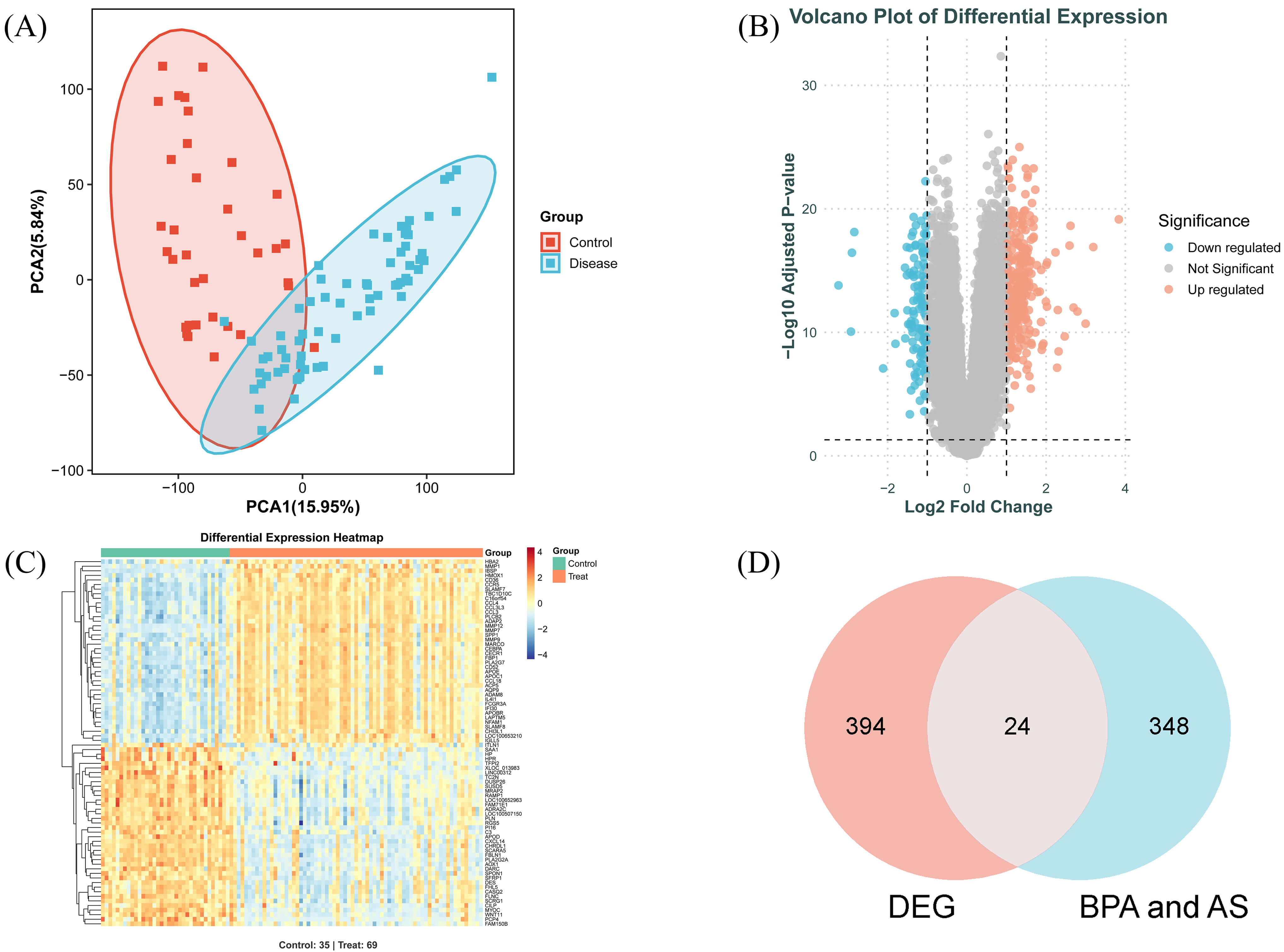 Fig. 3.
Fig. 3.
Analysis of DEGs and common target genes in AS. (A) PCA plot of the dataset. (B) Volcano plot of DEGs. Blue dots represent downregulated DEGs, red dots represent upregulated DEGs. (C) Heatmap of relative expression levels for selected DEGs. (D) Intersection Venn diagram. PCA, principal component analysis; DEGs, differentially expressed genes.
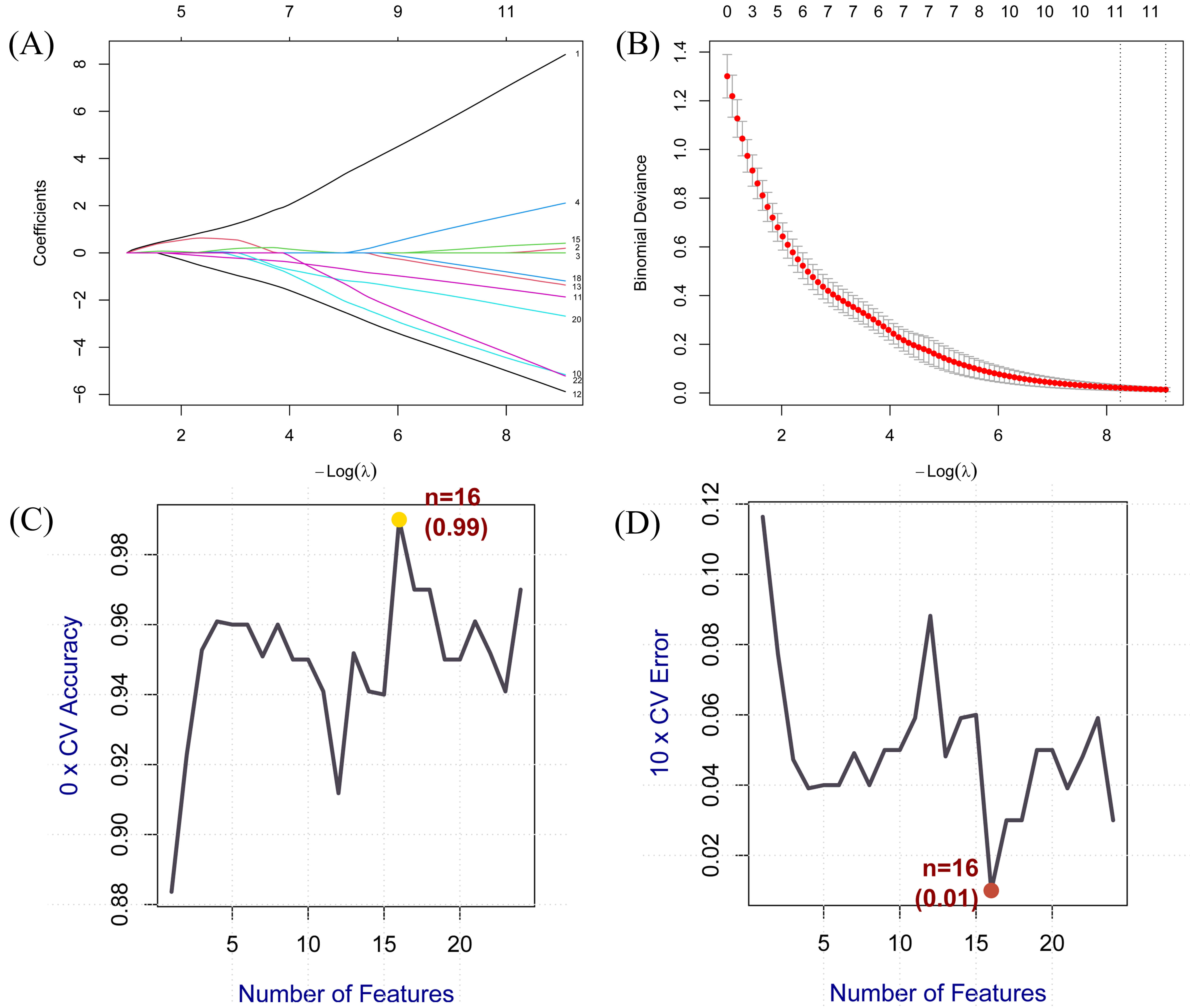
We employed LASSO and SVM algorithms to identify core genes from the DEGs across 24 common targets. LASSO regression optimised by 10-fold cross-validation and SVM-RFE identified 11 and 16 core genes respectively (Fig. 4).
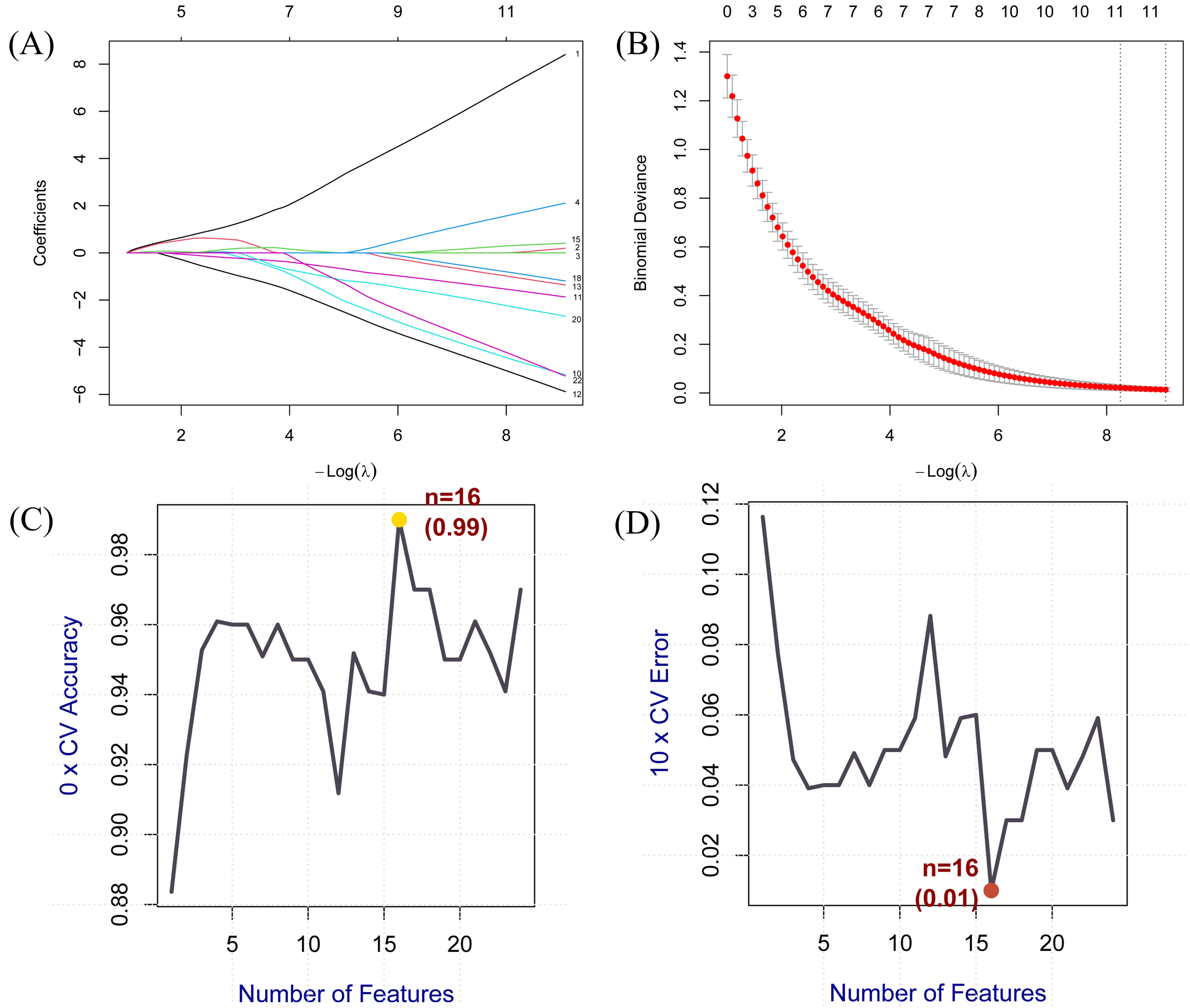 Fig. 4.
Fig. 4.
Core genes screened for BPA-induced AS using machine learning
algorithms. (A,B) Core genes identified via LASSO logistic regression. The
horizontal axis denotes the number of genes in the model corresponding to
different
By integrating core genes selected through CytoNCA, LASSO regression, and SVM-RFE, we identified TNF as a key core gene (Fig. 5A). To further validate TNF’s central role in AS, we constructed a ROC curve and measured the AUC to assess its diagnostic efficacy as a biomarker (Fig. 5B). ROC curve analysis revealed that TNF exhibited an AUC value approaching 1.0, indicating its ability to accurately distinguish AS lesion tissue from normal tissue and demonstrating strong diagnostic potential. Within the GSE100927 dataset, TNF expression was significantly upregulated in AS lesion tissue compared to normal controls, further highlighting its importance as a potential biomarker (Fig. 5C).
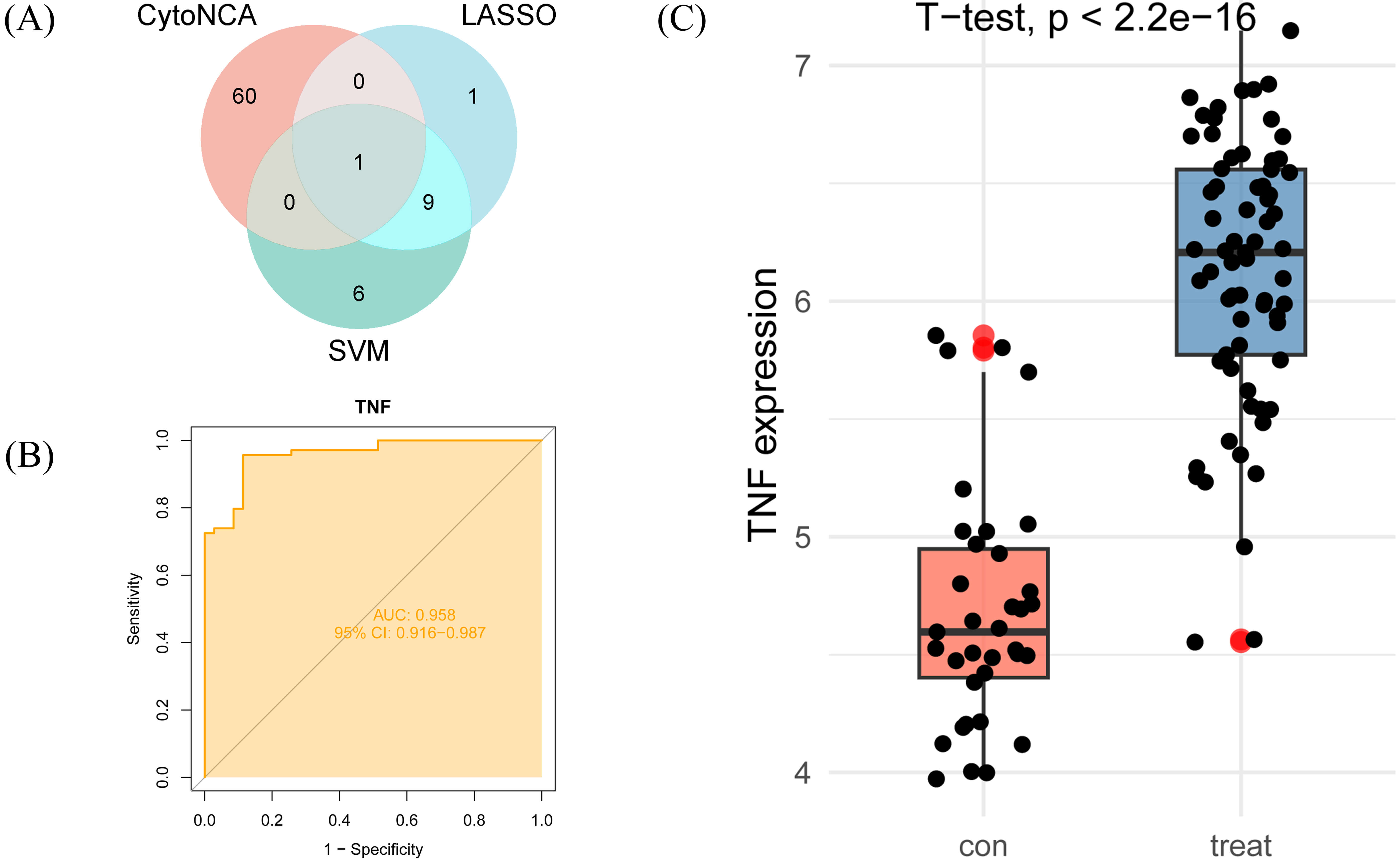 Fig. 5.
Fig. 5.
Identification of key core genes and validation of diagnostic efficacy. (A) A Venn diagram illustrating the intersection of genes selected by CytoNCA, LASSO regression, and SVM-RFE. (B) Evaluation of TNF’s diagnostic performance using a ROC curve. (C) TNF expression in AS tissue and healthy controls (GSE100927 dataset).
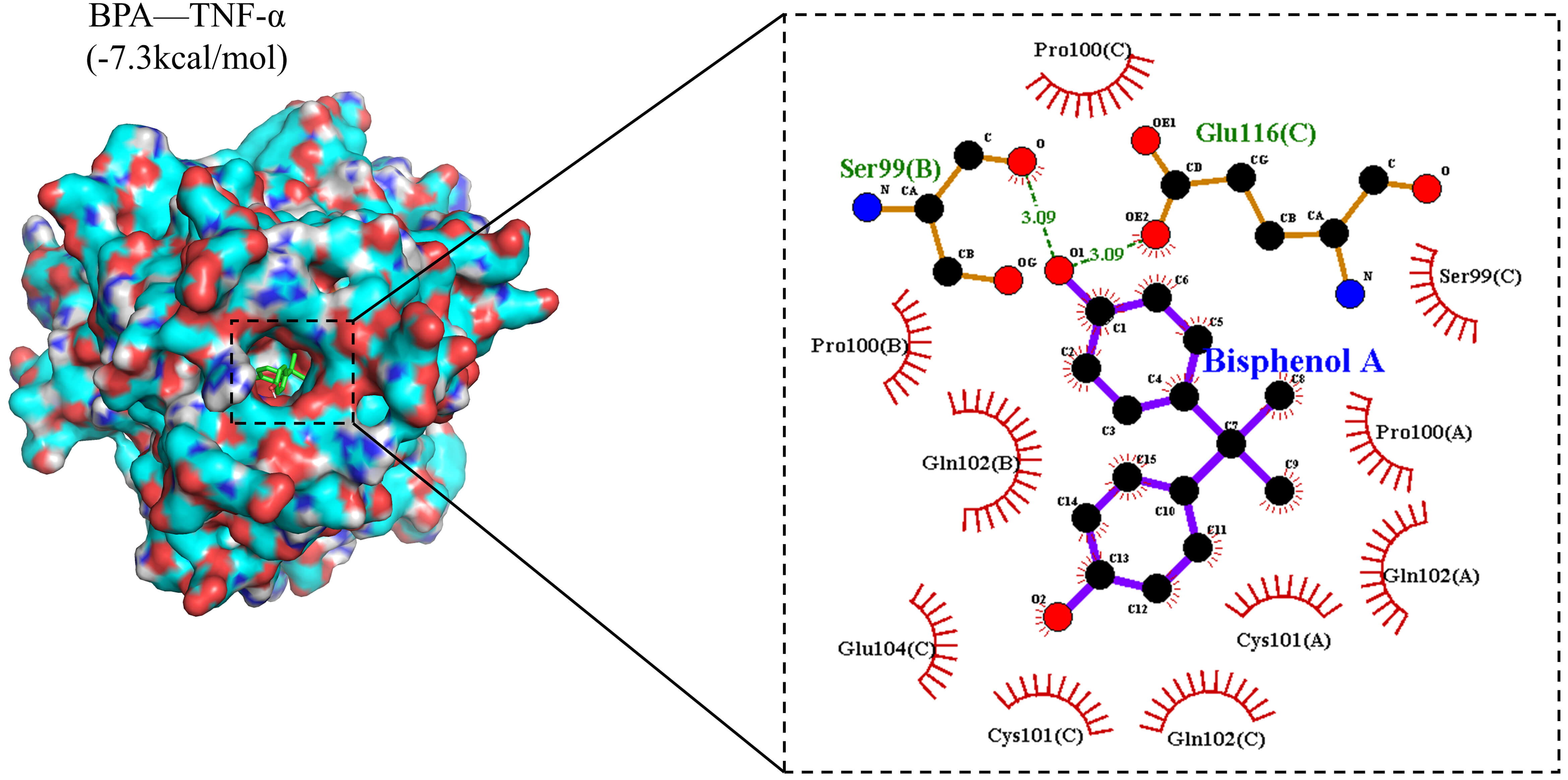
We employed molecular docking techniques to evaluate the binding between BPA and
the target protein tumour necrosis factor-
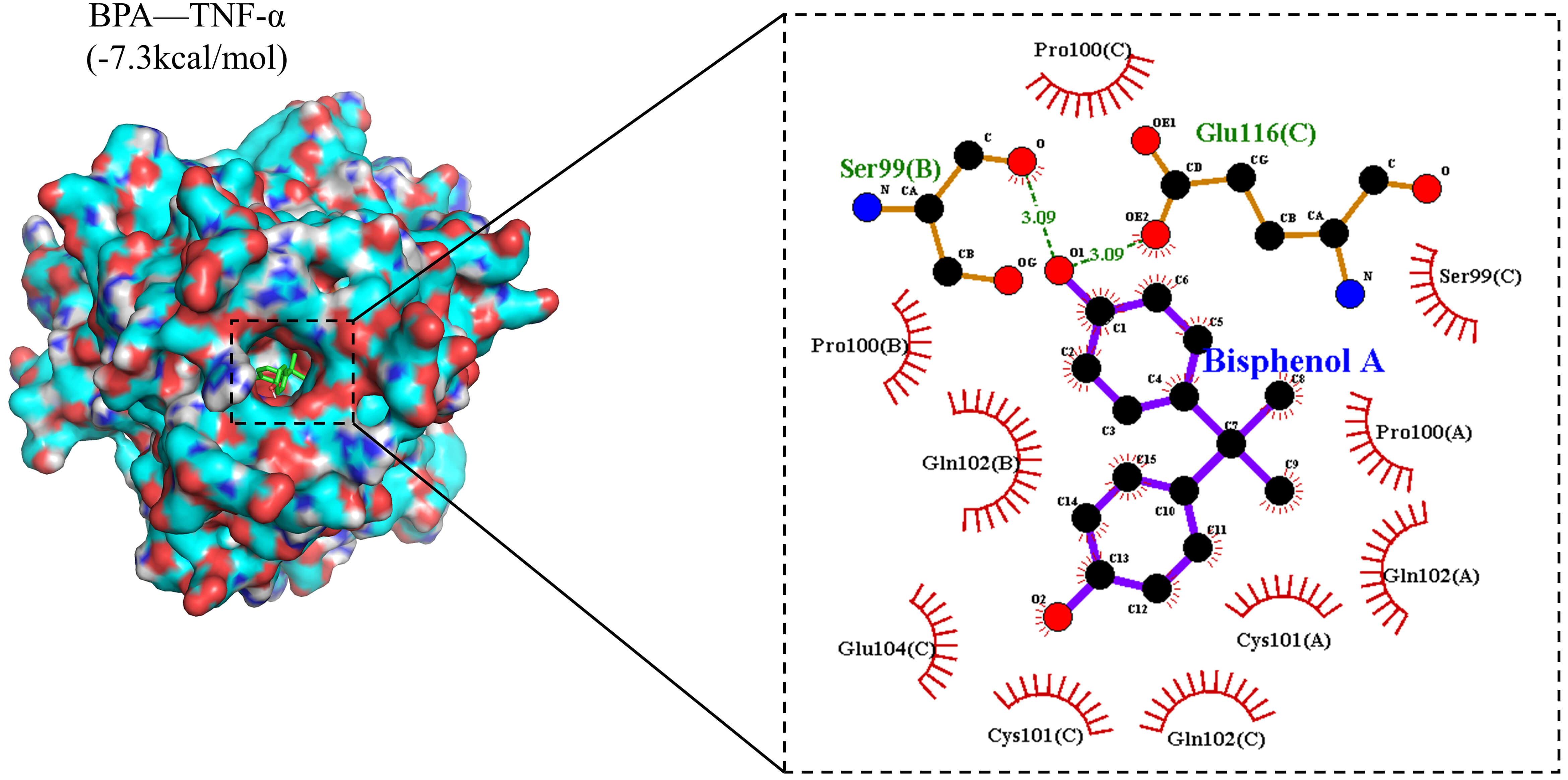 Fig. 6.
Fig. 6.
Two-dimensional (2D) and three-dimensional (3D) interaction diagrams of molecular docking results. In the two-dimensional interaction diagram, the purple ring-like structures represent wood-derived bisphenol A (BPA), the orange chain-like structures denote amino acids forming hydrogen bonds with BPA, the red arc-like structures indicate amino acids forming hydrophobic interactions with BPA, and the green dashed lines represent hydrogen bonds formed between BPA and the protein. Carbon atoms are displayed in black, oxygen atoms in red, nitrogen atoms in blue, and sulphur atoms in yellow.
This study systematically investigated the potential mechanisms linking BPA and AS by integrating multiple methodologies including ADMET analysis, network toxicology, transcriptomics, machine learning, and molecular docking. Findings indicate that BPA may contribute to the onset and progression of AS through multiple pathways, offering novel insights into the role of environmental pollutants in cardiovascular disease.
ADMET analysis revealed BPA’s significant biotoxicity and metabolic characteristics as an endocrine disruptor. BPA’s low intestinal absorption capacity, low plasma protein binding rate, and metabolic inhibitory effects suggest potential accumulation in vivo, particularly under chronic low-dose exposure, which may exacerbate AS progression. According to the European Food Safety Authority’s 2023 re-evaluation of BPA, a new tolerable daily intake of 0.2 ng/kg/day has been established, significantly lower than the previous provisional tolerable daily intake [39]. In contrast, the U.S. Environmental Protection Agency has historically published an oral reference dose for BPA of 0.05 mg/kg/day [40]. Notably, in vivo studies have provided evidence of BPA’s effects on AS. In an ApoE–/– mouse model fed a high-fat diet, continuous exposure to 50 µg/kg/day of BPA for 12 weeks significantly increased the burden of atherosclerotic lesions [41]. Furthermore, BPA has been widely detected in various environmental media. Literature reports indicate its concentration ranges from 16 to 1465 ng/L in industrial or domestic wastewater and from 170 to 3113 ng/L in surface water [42]. Against this backdrop, how BPA triggers inflammation-related alterations in the structure of the vascular wall at the molecular level is particularly crucial. And, consistent with prior studies, BPA’s interaction with NR-ER and SR-MMP receptors may further promote vascular remodelling and inflammatory responses, thereby driving AS development [43].
Through network toxicology analysis, we predicted 372 intersecting genes associated with BPA and AS from public databases, subsequently identifying 61 core genes via the CytoNCA plugin. KEGG enrichment analysis of these 372 intersecting genes revealed that these targets not only involve classical endocrine receptors but are also closely linked to multiple AS-related signalling pathways, including PI3K-Akt, MAPK, and Ras [44, 45]. These pathways play a crucial role in processes such as vascular endothelial function, smooth muscle cell proliferation, inflammatory responses, and lipid metabolism [46, 47, 48]. Specifically, the PI3K-Akt pathway is closely associated with the maintenance of endothelial cell function, promoting endothelial cell proliferation and repair while inhibiting endothelial dysfunction [49]. The MAPK pathway plays a pivotal role in the initiation and progression of inflammatory responses, particularly in the proliferation of smooth muscle cells and the formation of plaques [50]. The Ras signalling pathway regulates cell proliferation, migration and apoptosis, and is crucial for vascular remodelling and the progression of aortic stenosis [51]. Therefore, BPA may trigger a series of pathological changes in various cell types—such as endothelial cells, smooth muscle cells, and macrophages—through the interaction of these signalling pathways, thereby accelerating the development of aortic stenosis [52, 53, 54]. These findings are consistent with recent epidemiological and animal studies, with an increasing body of research suggesting that as an endocrine disruptor, BPA may contribute to the development of cardiovascular disease by modulating metabolic and immune systems [55, 56]. This study may point the way towards future approaches to mitigate or prevent BPA-induced vascular damage by targeting relevant signalling pathways.
To identify reliable gene targets, we screened 418 differentially expressed
genes (DEGs) from the GSE100927 dataset and intersected them with 372 genes,
ultimately yielding 24 common genes. Subsequently, using two complementary
machine learning algorithms—LASSO regression and SVM-RFE—we selected 11 and
16 high-confidence DEGs respectively. These genes were further intersected with
the CytoNCA-selected gene set, ultimately identifying TNF as the key core gene.
TNF-
To validate the pivotal role of TNF in AS, we constructed a ROC curve and
calculated the AUC value. The AUC value for TNF approached 1.0, indicating its
strong diagnostic potential and supporting the clinical application of TNF as a
biomarker for AS. Further molecular docking analysis validated BPA’s binding
affinity with TNF-
Although this study has thoroughly investigated the mechanisms by which BPA induces AS through approaches such as network toxicology, machine learning, and molecular docking, certain limitations remain. Firstly, while computational biology methods have predicted a potential binding relationship between BPA and TNF, the biological effects of this interaction have yet to be validated in in vitro or in vivo experiments. Consequently, future research should employ animal models or cellular experiments to further confirm BPA’s specific role in the TNF signalling pathway. Secondly, although TNF has been identified as a key core target, the development of AS represents a complex multifactorial, multi-pathway process. BPA may also influence AS progression via other targets. Consequently, future research should consider incorporating additional potential targets into analyses to provide a more comprehensive mechanistic explanation.
Beyond the TNF-centered mechanism of action highlighted in this study, it should
be noted that BPA may also influence the progression of AS through other
classical biochemical pathways, which have not been systematically investigated
in this research. For example, the eNOS/NO axis serves as a key endothelial
protective mechanism for maintaining vasodilation and inhibiting inflammatory
adhesion and thrombosis. Its dysfunction is considered a crucial molecular basis
for early endothelial injury in AS [61]. Previous study suggest that BPA exposure
may disrupt endothelial redox homeostasis, thereby affecting eNOS phosphorylation
status, enzyme activity, and NO bioavailability, and thus contribute to
endothelial dysfunction [62]. However, the causal relationship and molecular
mechanisms by which BPA acts on the eNOS/NO axis remain poorly understood and
require further investigation. Second, dysregulation of lipid homeostasis is a
core driver of AS plaque formation and progression [63]. Available evidence
indicates that low-dose BPA can upregulate the expression of cholesterol
synthesis-related genes through mechanisms such as reducing associated DNA
methylation, and exacerbates plaque burden and alters the lipid profile in AS
susceptibility models [64]. Additionally, BPA may downregulate apoA-I expression
by activating NF-
Based on existing research evidence, TNF-
This study successfully identified TNF as a key core gene in BPA-induced AS through integrated approaches including network toxicology, transcriptomics, machine learning, and molecular docking. Machine learning screening and molecular docking analysis further validated the direct binding between BPA and TNF, with the diagnostic potential of TNF as an AS biomarker assessed via ROC curves and AUC values. The findings indicate that TNF plays a pivotal role in the mechanism of BPA-induced AS and possesses strong diagnostic potential. This discovery provides a robust theoretical foundation for future applications of TNF as a clinical biomarker and therapeutic target.
AS, atherosclerosis; BPA, Bisphenol A; CVD, cardiovascular disease; LASSO, Least Absolute Shrinkage and Selection Operator; SVM-RFE, Support Vector Machine Recursive Feature Elimination; ROC, Receiver Operating Characteristic; AUC, Area Under the Curve.
All data generated or analyzed during this study are included in the published article [and its supplementary files], and are available from the corresponding author upon reasonable request.
Design and Concept: DBX and YWC; Literature Search and Screening: ZZ and MYZ; Data Analysis and Organisation: XX and XL; Graphic Production: JRH and SWX; Manuscript Drafting: All authors; The first draft was prepared by DBX. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
This research received support from Wenyi Deng from the Clinical Research Institute of the Second Affiliated Hospital of Hainan Medical University in data collation and map analysis. Special gratitude is also owed to Xiaozhi Lin for her encouragement and support.
This study was supported by the National Natural Science Foundation of China Regional Project (82360063) and the Hainan Province Graduate Innovation Research Project for General Higher Education Institutions (HYYB2024S002), as well as the Hainan Provincial Clinical Medical Center.
The authors declare no conflict of interest.
During the drafting of this article, the authors utilised ChatGPT-5 for spelling and grammar checks. Following the application of this tool, the authors undertook necessary review and editing of the content and assumes full responsibility for the published material.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/IJP48160.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.