

 , Zhongyang Deng 1,†, Hangrao Wang 2, Jie Zhou 2, Jinda Wang 2, Bei Jiang 1,2, Lei Shen 1,2,*
, Zhongyang Deng 1,†, Hangrao Wang 2, Jie Zhou 2, Jinda Wang 2, Bei Jiang 1,2, Lei Shen 1,2,*
1 Yunnan Key Laboratory of Screening and Research on Anti-pathogenic Plant Resources from Western Yunnan, Dali University, 671000 Dali, Yunnan, China
2 College of Pharmacy, Dali University, 671000 Dali, Yunnan, China
†These authors contributed equally.
Abstract
This study aimed to evaluate the therapeutic effect of Megacarpaea delavayi (M. delavayi) on L-arginine (L-arg)-induced functional dyspepsia (FD) and to elucidate the associated underlying mechanisms using network pharmacology and in vivo experiments.
A mouse model of L-arg-induced FD was established to assess the efficacy of M. delavayi. Subsequently, network pharmacology analysis was employed to predict potential candidate targets and signaling pathways of M. delavayi. Finally, the candidate targets were assessed in the mouse model described above.
M. delavayi promoted gastrointestinal peristalsis, reduced gastric tissue damage, increased serum levels of motilin (MTL) and gastrin (GAS), and decreased acetylcholinesterase (AChE) level in FD mice. Network pharmacology analysis identified 102 overlapping targets at the intersection of M. delavayi-related compounds and FD-associated genes. Protein-protein interaction (PPI) network analysis identified inflammation-associated targets, including interleukin-6 (IL6), tumor necrosis factor (TNF), nitric oxide synthase 1 (NOS1), epidermal growth factor receptor (EGFR), SRC proto-oncogene (SRC), and signal transducer and activator of transcription 3 (STAT3). Integration of these targets with functional and pathway enrichment analyses revealed that nitric oxide (NO) and related inflammatory signaling pathways play a pivotal role in the pharmacological effects of M. delavayi. Validation in FD mice further demonstrated that M. delavayi decreased O₂⁻ contents and restored the impaired NO-soluble guanylate cyclase (sGC)-protein kinase G (PKG) signaling pathway by elevating the levels of neuronal nitric oxide synthase (nNOS), sGC and PKG. Conversely, an increase in inducible nitric oxide synthase (iNOS) expression was also observed.
M. delavayi effectively ameliorates L-arg-induced gastrointestinal dysfunction through restoration of the NO-sGC-PKG signaling pathway.
Keywords
- Megacarpaea delavayi
- functional dyspepsia
- nitric oxide
- soluble guanylate cyclase
- protein kinase G
- network pharmacology
Functional dyspepsia (FD) is a prevalent, chronic, and recurrent gastrointestinal disorder with an estimated prevalence ranging from 10% to 40% in Western countries and 5% to 30% in Asia [1]. It is characterized by symptoms such as epigastric discomfort, bloating, early satiety, belching, nausea, and acid regurgitation, in the absence of detectable organic pathology. Although FD is not a life‑threatening disease, it markedly impairs work performance and quality of life, and places a considerable economic burden on individuals and healthcare systems. Current medicines for FD include proton pump inhibitors, prokinetic agents (e.g., domperidone and metoclopramide), antidepressants, and mucosal protectants [2, 3]. However, their clinical applications are limited by potential adverse effects, variable efficacy among individuals, and high relapse rates following discontinuation [4, 5].
Ethnomedicines are guided by traditional medical theories and supported by long-term clinical practice. Therefore, their application in treating chronic refractory diseases such as FD represents a feasible therapeutic strategy [6]. In Dali, Yunnan province of China, a plant, Megacarpaea delavayi (M. delavayi) Franchet, has been employed as a traditional ethnic medicine for centuries to treat FD. M. delavayi, commonly known as Gaohecai (Chinese name) or Ganlgect (Bai ethnic name), is a perennial herb that belongs to the genus Megacarpaea in the Brassicaceae family. It grows in high-altitude regions at elevations between 3400 and 3800 meters. This genus comprises seven species worldwide, three of which are distributed in China, mainly in Yunnan, Sichuan, Gansu, and Tibet. According to traditional medicinal records of the Bai ethnic group, M. delavayi is used as both a medicinal and a food plant for treating dysentery, gastric heat accumulation, indigestion, and heat-related cough. Recent pharmacological studies have demonstrated that M. delavayi can promote gastric emptying and small intestinal propulsion in both normal mice and those with FD, enhance contractions of the isolated guinea pig ileum, and stimulate gastric acid secretion in rats with food stagnation [7, 8, 9]. However, its comprehensive therapeutic efficacy and underlying mechanisms remain insufficiently elucidated.
Although the pathophysiology of FD remains incompletely understood, several key mechanisms have been proposed, including disordered gastrointestinal motility associated with central modulation, visceral hypersensitivity to mechanical or chemical stimuli, dysregulation of the brain-gut axis, and Helicobacter pylori infection [10]. Recently, increasing evidence has revealed the role of nitric oxide (NO) and its related signaling pathway in the regulation of gastrointestinal function. In the gastrointestinal tract, NO is an essential inhibitory neurotransmitter that is biosynthesized through the catalysis of nitric oxide synthase (NOS). Physiologically, NO generated by endothelial NOS (eNOS) or neuronal NOS (nNOS) modulates smooth muscle relaxation via the NO-guanylate cyclase (GC)-protein kinase G (PKG) pathway [11, 12, 13]. However, under FD conditions, both the impaired NO synthesis from nNOS dysfunction and abnormal NO release mediated by inducible NOS (iNOS) are closely associated with gastrointestinal dysmotility. Dysregulation of the NO–sGC–PKG signaling pathway may lead to delayed gastric emptying and impaired accommodation, which are key features of FD [14, 15, 16, 17]. Excessive intake of L-arginine (L-arg), a metabolic precursor of NO, could lead to overproduction of NO both directly and indirectly, and could further impair gastrointestinal function [18]. Compared with other classical FD animal models, such as those induced by iodoacetamide or tail clamp stimulation, the L-arg model can successfully replicate key clinical features of FD, including delayed gastric emptying and reduced intestinal propulsion, without causing obvious necrosis, erosion, or organic lesions. Thus, this model can closely mimic human FD [19, 20, 21].
This study investigated the efficacy of M. delavayi in treating FD and explored its underlying mechanism in an L-arg-induced mouse model. These findings may provide a scientific basis for the traditional application of M. delavayi in FD treatment.
Male SPF grade ICR mice (18–22 g) were procured from Speforward (Beijing) Biotechnology Co., Ltd. (License No. SCXK (Beijing) 2024-0001). Mice were housed under standard laboratory conditions (18–22 °C, 50–60% relative humidity) with a 12 h light/dark cycle and had ad libitum access to food and water. Animals were acclimatized to the laboratory environment for one week prior to experimentation. All animal procedures were performed in accordance with the guidelines of the Animal Ethics Committee of Dali University (Ethics No. 2024-P2-110).
The whole plant of M. delavayi was collected in Dali City, Yunnan Province, China, and authenticated morphologically by Dr. Yong-Zeng Zhang at the College of Pharmacy, Dali University, Yunnan Province, China. A voucher specimen was deposited at the Yunnan Key Laboratory of Screening and Research on Anti-pathogenic Plant Resources from Western Yunnan, Dali University (plant sample preservation number: 20180627-4). Two kilograms of plant samples were milled and extracted three times with 95% ethanol at room temperature (24 h, 48 h, and 48 h, respectively). The resulting extracts were combined, filtered, and concentrated under reduced pressure using a rotary evaporator, yielding 445 g of freeze-dried powder. Prior to administration, the powders were suspended in a 0.5% CMC-Na solution. CMC-Na (Batch No: IS9000) was purchased from Solarbio Biotechnology Co., Ltd., Beijing, China. Domperidone was purchased from Shanghai Yuan-Ye Co., Ltd. (Batch No: S26612) and used as a positive control drug.
L-arg (Batch No: 150905) and superoxide anion (O₂⁻) detection kit (Batch No:
BC1290) were purchased from Solarbio Biotechnology Co., Ltd., China. Evans Blue
dye (Batch No. 991107) was purchased from the Shanghai Chemical Reagents
Procurement and Supply Station. Detection kits of motilin (MTL, Batch No:
H182-1-2) and gastrin (GAS, Batch No: H239-1-2) were sourced from Nanjing
Jiancheng Bioengineering Co., Ltd., China. Detection kits of vasoactive
intestinal peptide (VIP, Batch No: E-EL-M1234) and acetylcholinesterase (AChE,
Batch No: E-EL-M26) were purchased from Elabscience Biotechnology Co., Ltd.,
China. Detection kits of soluble guanylate cyclase (sGC, Batch No: ml925127V),
protein kinase G (PKG, Batch No: ml037829V), inducible nitric oxide synthase
(iNOS, Batch No: ml057773V), and neuronal nitric oxide synthase (nNOS, Batch No:
ml0930900) were sourced from Shanghai Enzyme-linked Biotechnology Co., Ltd.,
China. The primary antibodies against sGC (Batch No: 19011-1-AP), PKG (Batch No:
21646-1-AP), nNOS (Batch No: 29231-1-AP), iNOS (Batch No: 22226-1-AP), and
After acclimation, sixty mice were randomly assigned to six groups (n = 10 per group): control, model, domperidone (0.1 g/kg, positive control), and M. delavayi extract (1.0, 0.5, and 0.25 g/kg). All treatments were administered once daily by gavage for 7 days at an administration volume of 20 mL/kg. From days 4–7, all mice except those in the control group received L-arg (3 g/kg, 1 h before drug treatment) intragastrically to induce FD. The dosage and administration period were determined based on preliminary experiments and previously published studies.
A blue-colored semi-solid nutritional paste (300 mL) was prepared by dissolving
10 g sodium carboxymethyl cellulose in 250 mL distilled water, followed by
sequential addition of 12 g sucrose, 16 g milk powder, and 10 g starch to form a
suspension under continuous stirring, with the final addition of 0.1 g Evans Blue
dye as a visual tracer. The paste was stored at 4 °C and warmed to room
temperature prior to oral administration. After seven days of administration,
mice were for fasted 12 h with ad libitum access to water. The next morning, each
mouse was administered 0.8 mL of the prepared paste through gavage. Twenty
minutes post-gavage, the blood of each mouse was collected. Following collection,
mice were euthanized by cervical dislocation, and their abdomen was excised.
After ligation of both the cardia and pylorus, the entire stomach was removed and
rinsed with saline to eliminate blood contaminants. The total weight of stomach
was weighed, then the stomach was opened along the greater curvature, and the
gastric contents were washed with saline. The emptied stomach was wiped dry with
filter paper before measurement of the net weight of the stomach. Gastric
emptying rate (%) was calculated as: [Weight of administered paste – (total
weight of stomach – net weight of stomach)] / Weight of administered paste
Blood samples were collected from the retro-orbital plexus under brief isoflurane (3% for induction and 1.5–2% for maintenance, administered via inhalation using a calibrated vaporizer with oxygen flow) anesthesia and allowed to stay at room temperature for 2 h. Then, serum was prepared by centrifugation (3000 rpm for 10 min) at 4 °C, stored at –80 °C, and used to analyze the levels of gastrointestinal hormone. The gastric antrum specimen was isolated from the stomach and fixed in 4% paraformaldehyde for histopathological examination. The ileum tissue was obtained from the small intestine and homogenised on ice for subsequent analysis, including Enzyme-Linked Immunosorbent Assay (ELISA) and WB.
Following fixation, gastric specimens were processed through standard H&E staining protocols, including paraffin embedding, sectioning, xylene dewaxing, graded ethanol rehydration, H&E staining, ethanol dehydration, xylene clearing, and neutral balsam mounting. Histopathological changes were observed under a light microscope.
ELISA kits were employed to quantify the levels of gastrointestinal hormones and the contents of the key NO-sGC-PKG signaling pathway components in mice. After the preparation of serum, the contents of MTL, GAS, VIP, and AChE were measured according to the kit protocols. The samples of ileum tissues were homogenized and centrifuged, and the resulting supernatant was used to measure the contents of sGC, PKG, nNOS, iNOS and O₂⁻ with the kit instructions.
Ileal tissues were lysed in high-efficiency RIPA lysis buffer (Solarbio,
Beijing, China; Batch No. 20220211) supplemented with a 50
A total of 49 compounds isolated from the 95% ethanol extract of M.
delavayi (as reported by Tang [22]) were subjected to network pharmacology
analysis. Potential molecular targets of the identified compounds were predicted
using the SwissTargetPrediction database (http://www.swisstargetprediction.ch/),
and targets with a probability
Data were presented as mean
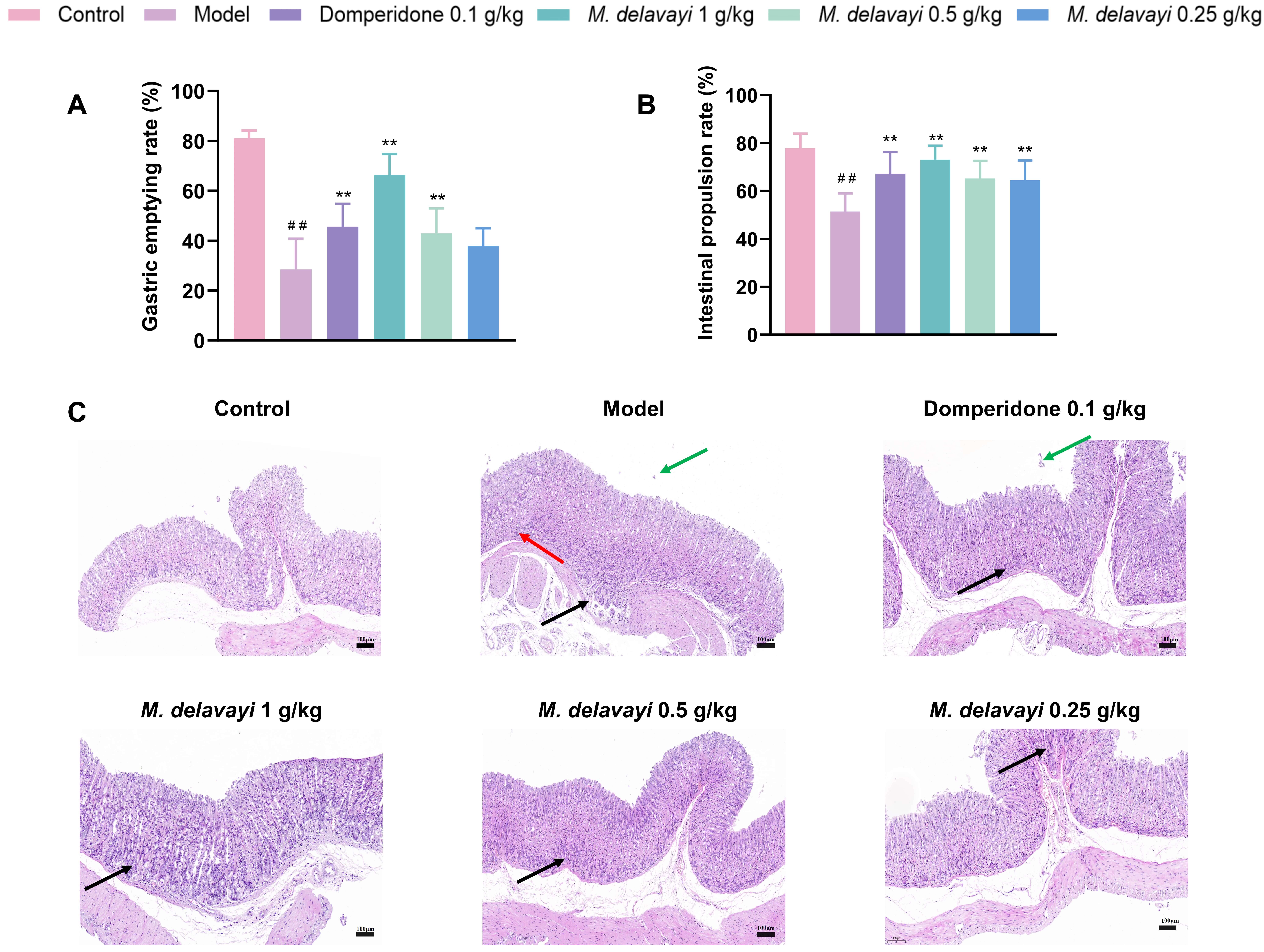
Compared to the control group, L-arg administration significantly reduced
gastric emptying rate and intestinal propulsion rate, both of which are key
indicators of abnormal gastrointestinal motility observed in the model group
(p
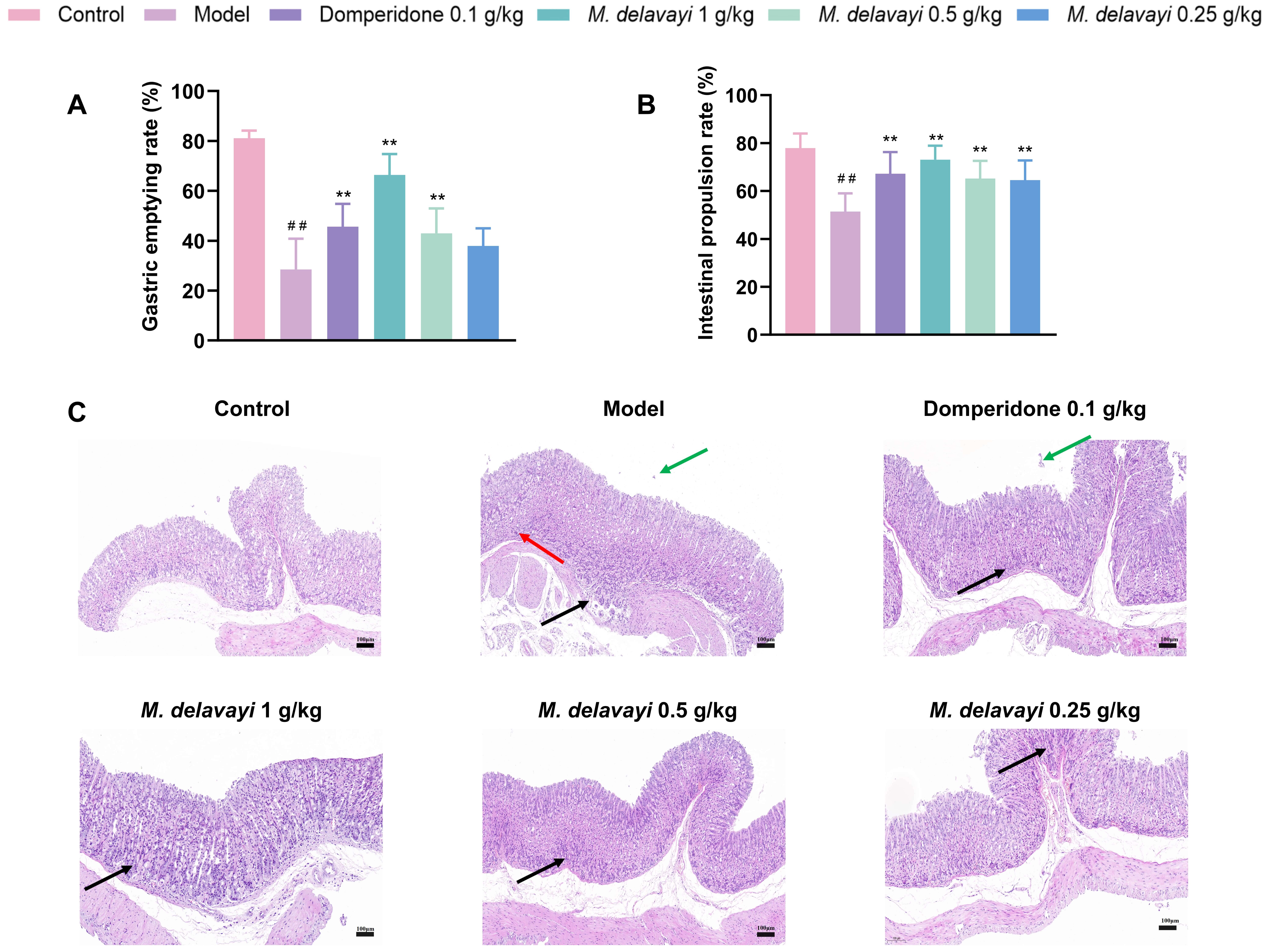 Fig. 1.
Fig. 1.
Effect of M. delavayi on gastrointestinal motility and
gastric tissue damage in FD mice. (A) Gastric emptying rate (n = 10). (B) Small
intestinal propulsion rate (n = 10). ## p
In the control group, H&E staining of the gastric antrum tissue exhibited uniformly stained cells and orderly arranged muscle fibers in the muscularis layer. The lamina propria contained abundant gastric glands with intact morphology. There were no obvious pathological alterations or inflammatory cell infiltration observed (Fig. 1C). In contrast, the model group displayed gastric tissue abnormalities, including desquamation of epithelial cells (green arrows) and focal necrosis of gastric glandular cells (red arrows, pyknotic nuclei) in the lamina propria, and marked infiltration of inflammatory cells (black arrows) in the submucosal layer (Fig. 1C). After domperidone and M. delavayi treatment, gastric antrum tissue exhibited varying degrees of improvement in structure, with relatively preserved epithelial integrity and reduced inflammatory infiltration (Fig. 1C).
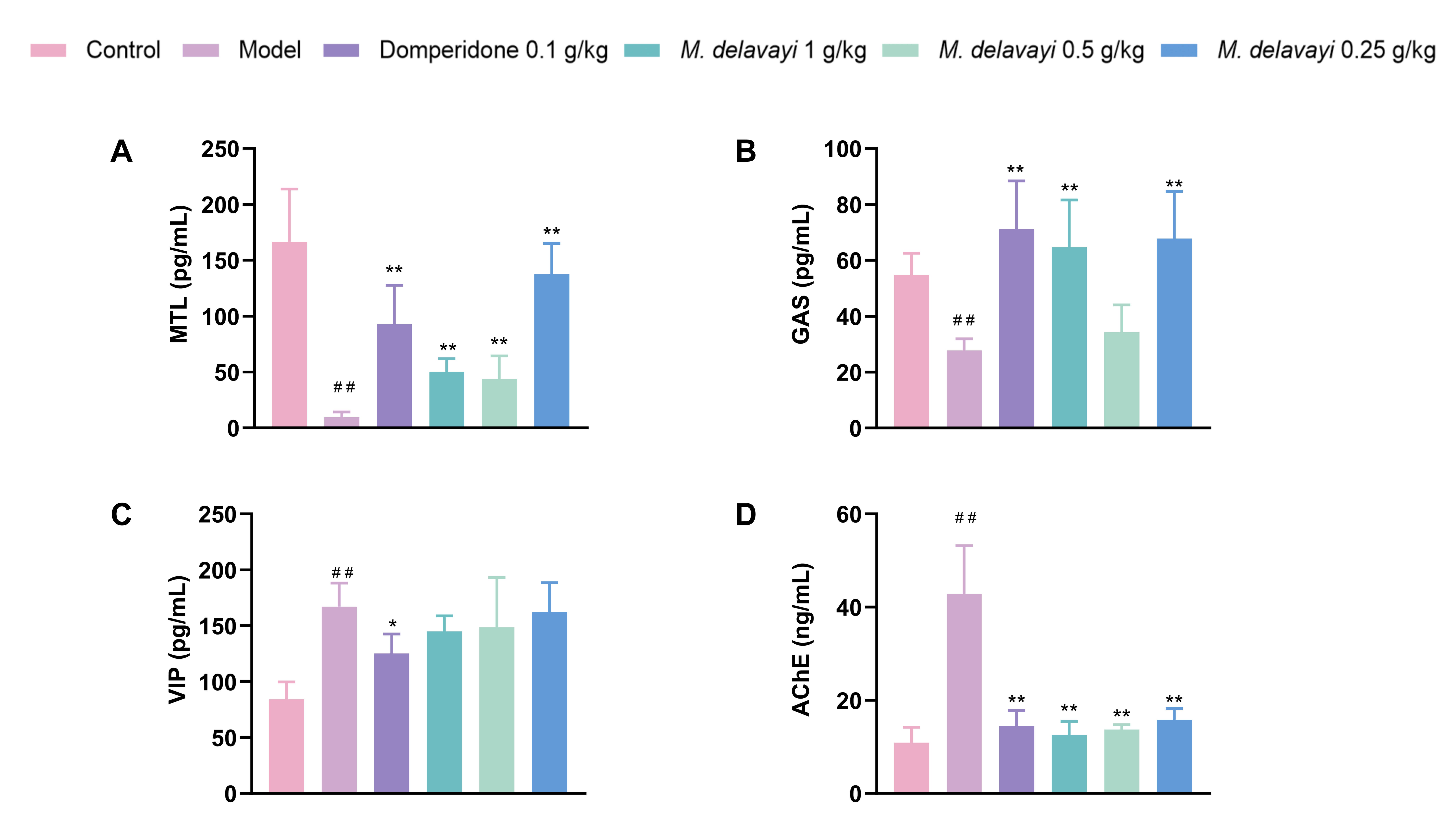
Gastrointestinal hormones play important roles in regulating gastric motility
and peristaltic rhythm. Compared to the control group, serum levels of motilin
MTL and GAS were significantly reduced in the model group (p
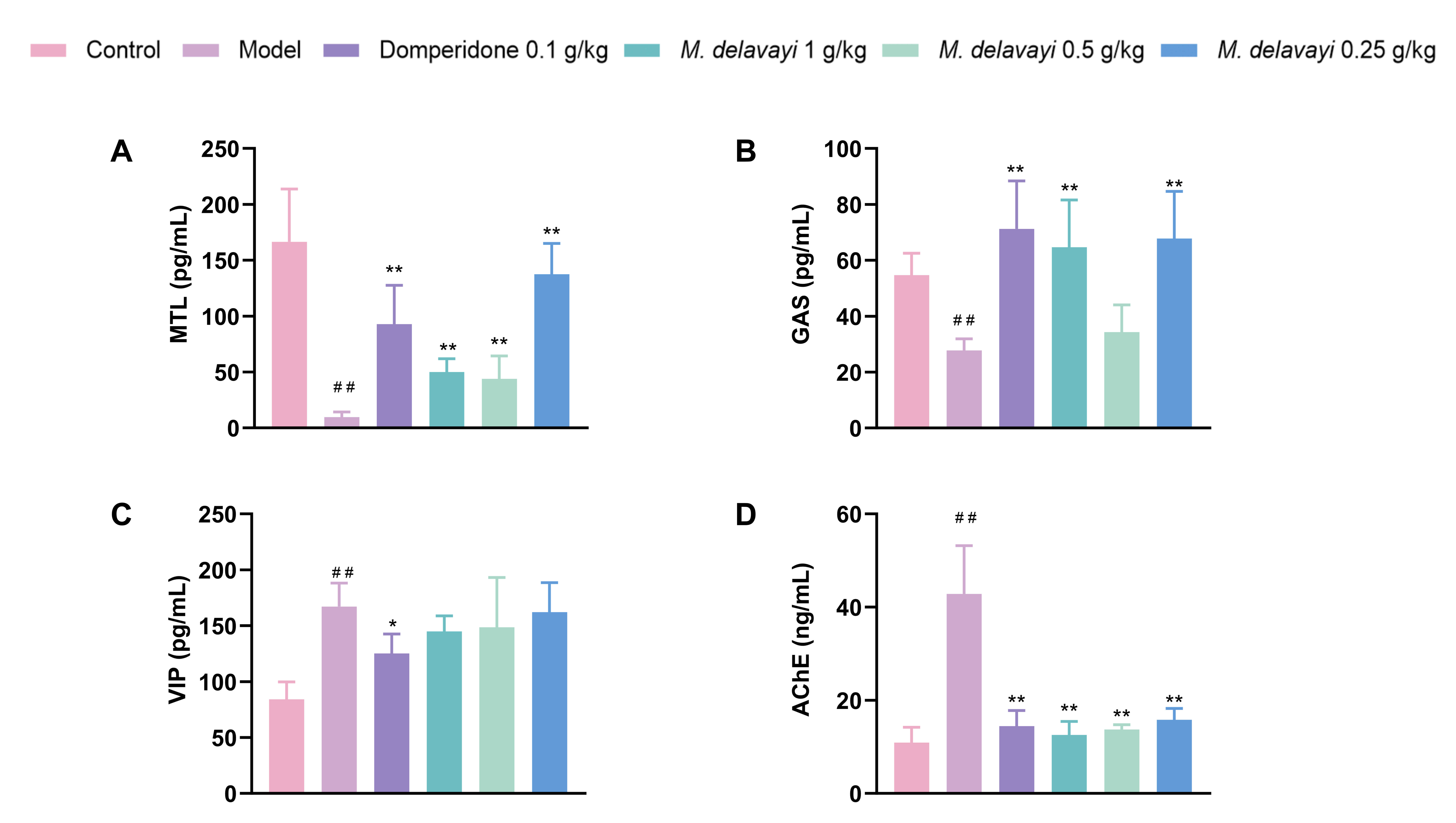 Fig. 2.
Fig. 2.
Effect of M. delavayi on serum levels of
gastrointestinal hormones and AChE in FD mice (n = 6). (A) MTL level. (B) GAS level. (C)
VIP level. (D) AChE level. ## p
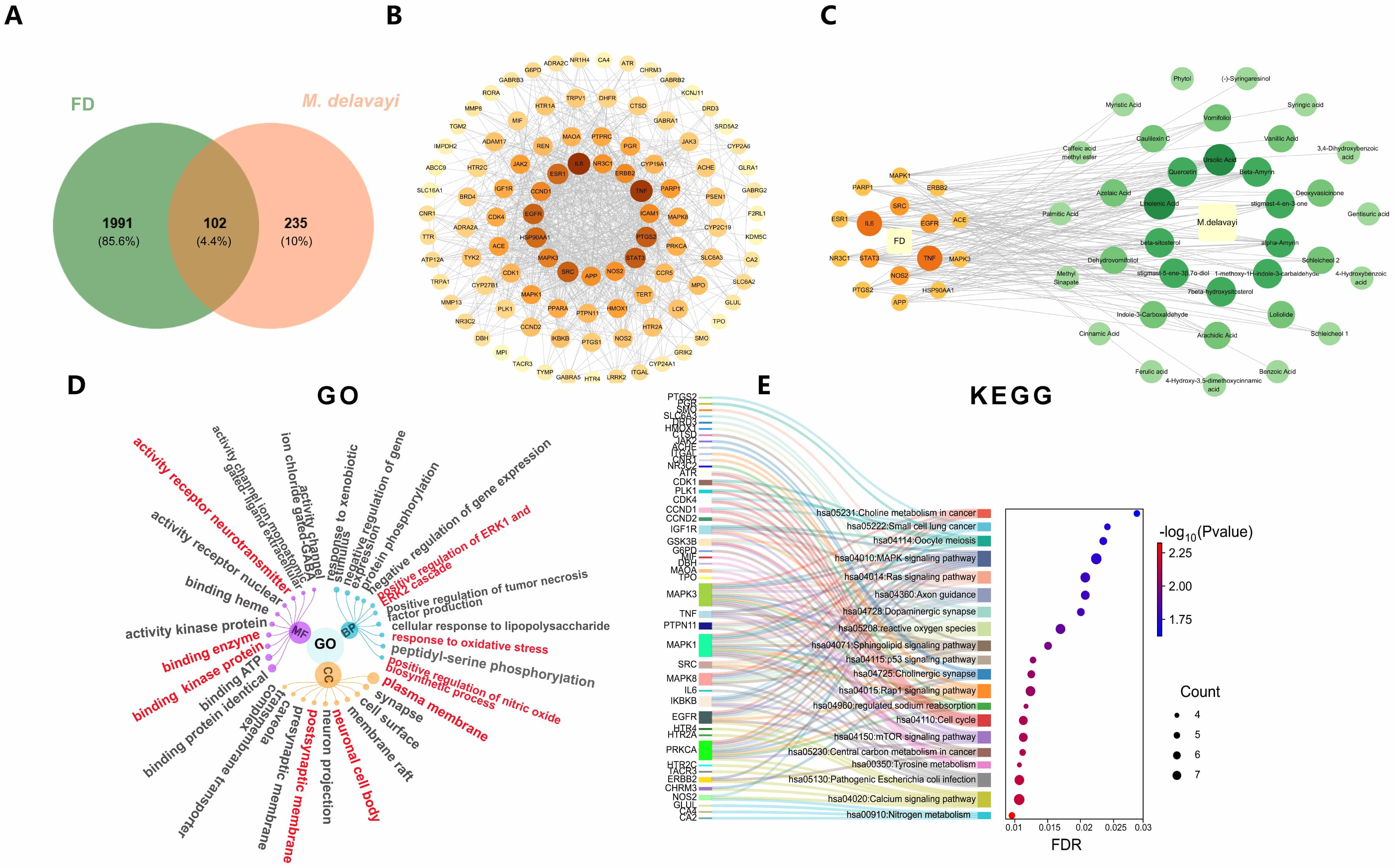
Based on previous phytochemical investigations, 49 active compounds were identified from M. delavayi. Among these, 35 compounds (Supplementary Table 1) were predicted to have corresponding protein targets. After target prediction using the SwissTargetPrediction database and removal of duplicate targets, 337 potential protein targets were obtained. Meanwhile, 2093 FD-associated genes were retrieved from three databases. Comparative analysis of compound-related targets and disease-associated genes yielded 102 overlapping targets, which were visualized in a Venn diagram (Fig. 3A).
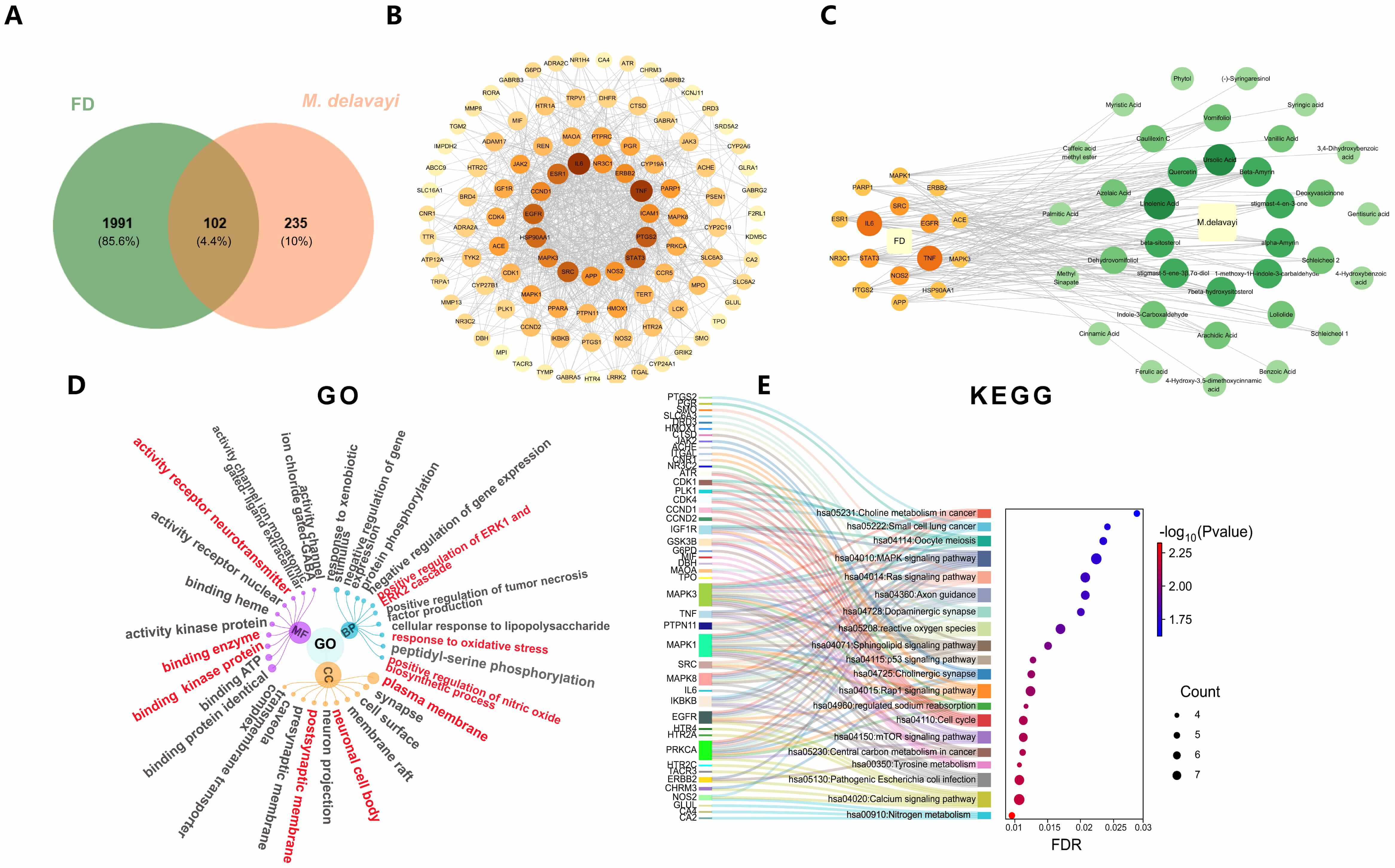 Fig. 3.
Fig. 3.
Network pharmacology prediction. (A) Venn diagram of overlapping target genes between FD and M. delavayi. (B) PPI network visualization of the total of 102 overlapping target genes. (C) PPI network visualization of the top 16 targets and their corresponding compounds in M. delavayi. (D) Hierarchical network plot for GO enrichment analysis (top 10 items). (E) Sankey diagram (left) and bubble plot (right) for KEGG enrichment analysis (top 20 items). BP, biological process; MF, molecular function; CC, cellular component; KEGG, Kyoto Encyclopedia of Genes and Genomes.
The PPI network of the 102 overlapping targets was analyzed using the STRING database and constructed using Cytoscape software. The network consisted of 102 nodes and 847 edges, with an average node degree of 6.48 (Fig. 3B). These targets were further filtered and explored through PPI network analysis, leading to the identification of 16 core targets. Among these, six are associated with inflammation, including IL6, TNF, NOS1, EGFR, SRC, and STAT3 (Fig. 3C).
GO enrichment analysis was performed across the categories of biological process
(BP), molecular function (MF), and cellular component (CC), using a significance
threshold of p
KEGG pathway enrichment analysis identified 120 enriched entries, and the top 20 significantly enriched signaling pathways were visualized using a bubble plot (Fig. 3E). The results demonstrated that the nitrogen metabolism, calcium signaling pathway, cholinergic synapse, and reactive oxygen species (ROS) were the main enriched pathways.
Integration of the core targets with GO and KEGG enrichment analysis revealed that NO and its associated inflammatory signaling pathways play a pivotal role in the action of M. delavayi, requiring further validation.
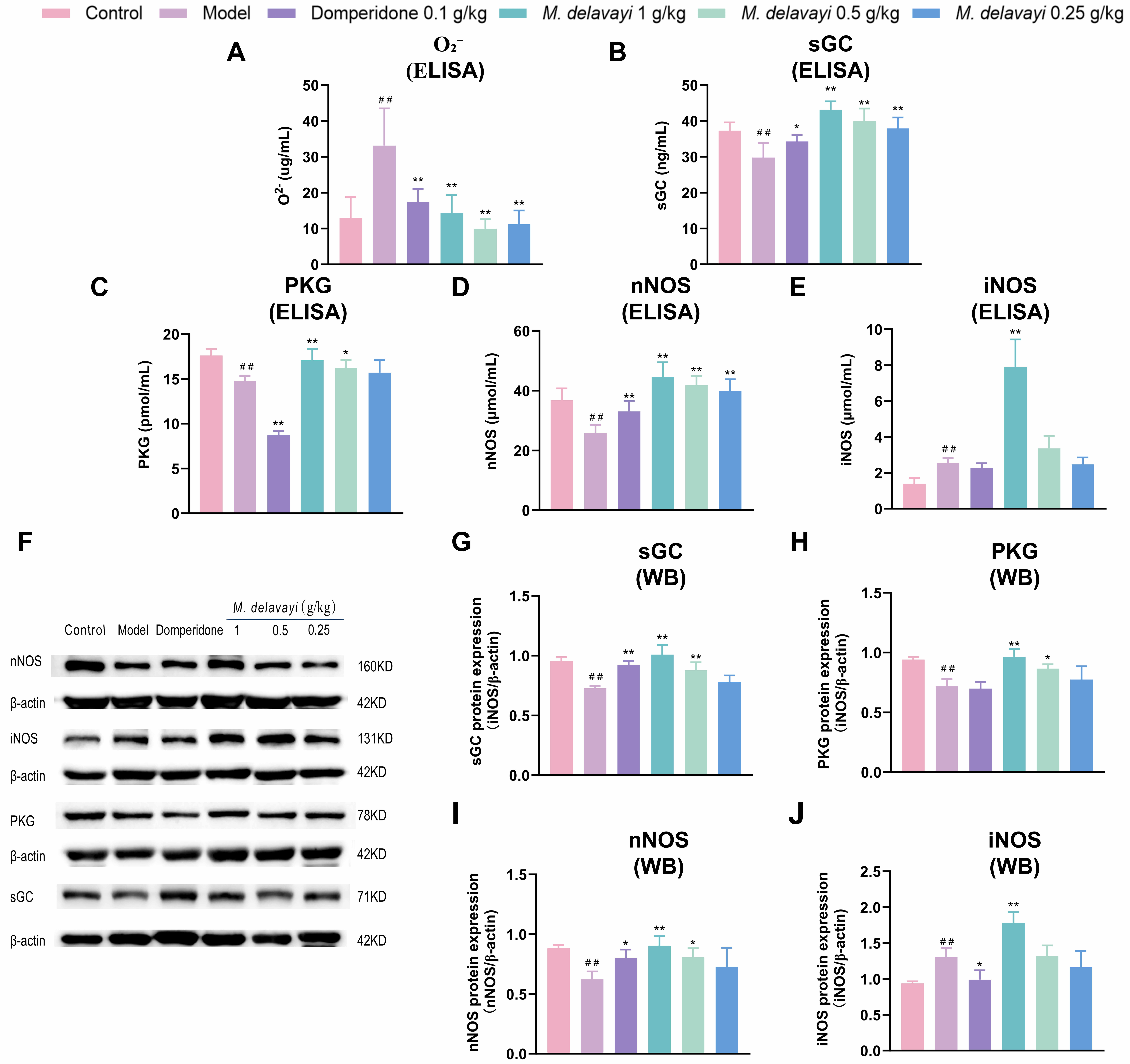
Firstly, the ELISA assay was used to quantify the levels of key components in
the NO-sGC-PKG signaling pathway, which is closely associated with
neurotransmitter receptor modulation, inflammatory responses, and oxidative
stress regulation. Compared to the normal group, L-arg intervention in FD mice
significantly elevated O₂⁻ contents (p
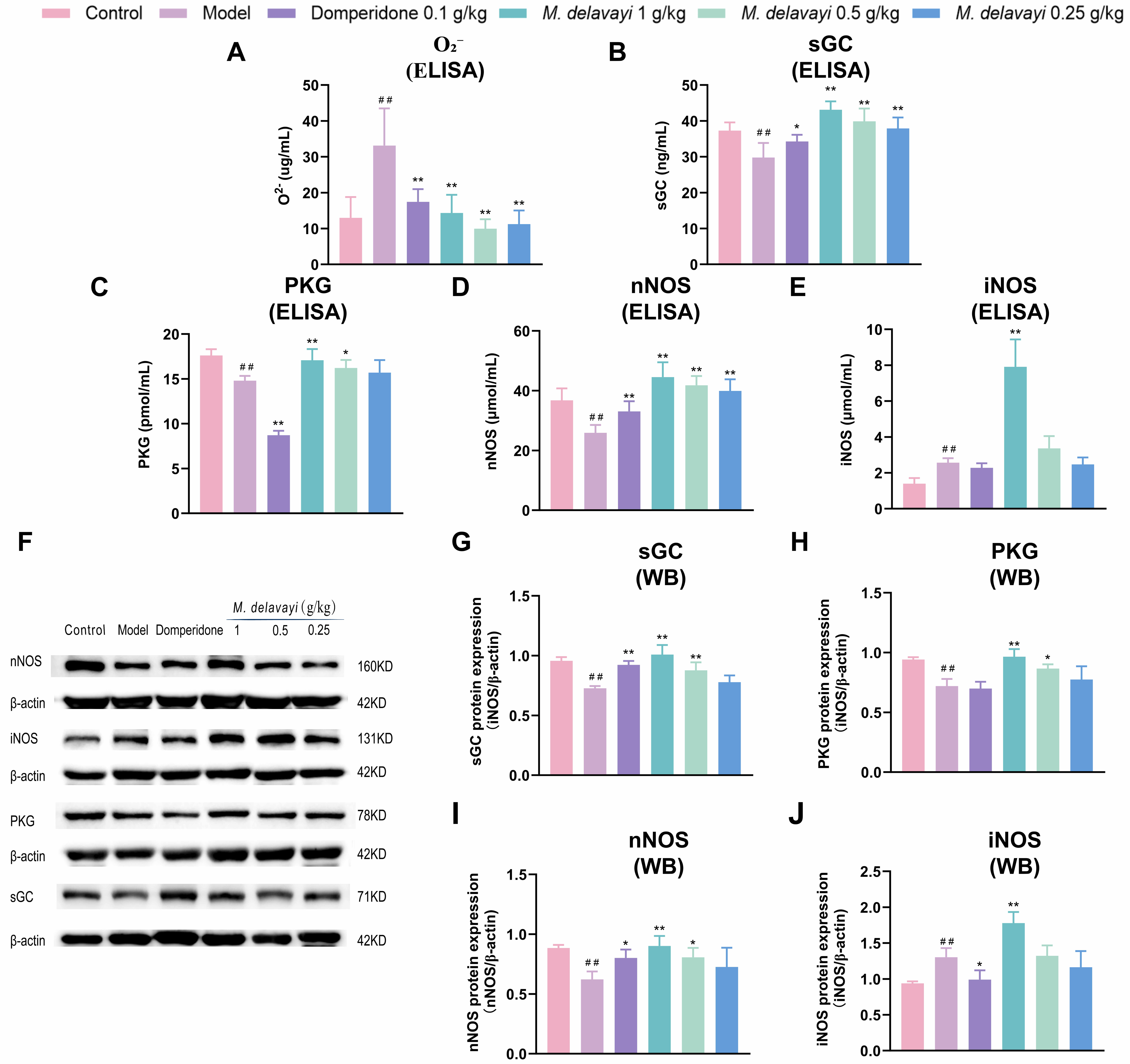 Fig. 4.
Fig. 4.
Effect of M. delavayi on the NO-sGC-PKG signaling
pathway in FD mice. (A–E) The levels of O₂⁻, sGC, PKG, nNOS, and iNOS were
measured by ELISA (n = 8). (F) Western Blot image. (G–J) The relative protein
levels of sGC, PKG, nNOS, and iNOS were measured by Western Blot (n = 4).
## p
Similar results were observed by WB analysis. Compared to the model group, 1 and
0.5 g/kg M. delavayi significantly increased the protein expression
levels of sGC, PKG, and nNOS (p
As a traditional medicine and food plant, M. delavayi has long been used by the Bai people in Dali of China to treat FD. However, modern pharmacological evidence supporting its efficacy is still inadequate. Here, our results demonstrate that M. delavayi effectively improves gastrointestinal motility in an L-arg-induced FD mouse model, which closely mimics the human disease. These findings are consistent with the ethnomedicinal use of M. delavayi by local populations, where it has been consumed to enhance appetite, strengthen the spleen, and relieve indigestion, thus providing experimental support for its traditional application.
Gastrointestinal motility is modulated by the autonomic and enteric nervous systems (ANS and ENS), as well as various gastrointestinal hormones, including MTL, GAS, and VIP. MTL and GAS promote gastric emptying, gastric acid secretion, and mucosal growth, whereas VIP induces smooth muscle relaxation via vasoactive intestinal polypeptide receptor activation and myosin light chain kinase inhibition [23, 24]. These hormones function in a coordinated and dynamic manner to fine-tune gastric accommodation, peristaltic rhythm, and pyloric relaxation, thereby maintaining physiological gastrointestinal motility. Disruption of neural regulation and gastrointestinal hormone secretion can lead to FD. In the L-arg-induced FD model, reduced levels of MTL and GAS contribute to delayed gastric emptying, whereas impaired NO-mediated inhibitory signaling may trigger compensatory upregulation of VIP by the ENS to maintain smooth muscle relaxation and prevent hypercontractility [25]. Such compensatory hormonal alterations reflect an adaptive response of the ENS to disturbed nitrergic signaling; however, persistent imbalance may further aggravate dysmotility and impair coordinated gastrointestinal movement. Our findings indicate that M. delavayi extract restores gastrointestinal function by elevating the levels of MTL and GAS and reducing abnormally elevated VIP level, thereby improving gastric emptying, intestinal propulsion, and motility rhythm. In addition to hormonal regulation, cholinergic signaling plays a crucial role in smooth muscle activity, where ACh mediates muscular contraction via the M3 receptor [26]. In the FD model, excessive L-arg intake disrupts physiological NO signaling and is associated with reduced nNOS activity, thereby impairing normal NO synthesis [27]. This diminishment of inhibitory tone weakens the control of cholinergic neurons, ultimately eliciting compensatory cholinergic hyperactivity characterized by increased ACh release and upregulated AChE activity [28, 29]. Here, M. delavayi alleviates these cholinergic abnormalities by inhibiting AChE overactivity. Collectively, these results suggest that M. delavayi exerts multiple regulatory effects on neurohormonal and neurotransmitter pathways, providing mechanistic insight into its therapeutic potential for FD.
Network pharmacology was utilized to systematically identify bioactive
components, therapeutic targets, and enriched pathways associated with M.
delavayi. GO and KEGG enrichment analyses revealed strong associations with NO
biosynthesis, oxidative stress regulation, inflammatory signaling, and calcium
homeostasis. Physiologically, NO produced by NOS diffuses into smooth muscle or
interstitial cells of Cajal, activates sGC, increases cGMP, and stimulates PKG,
thereby reducing intracellular Ca2⁺ levels and suppressing pro-inflammatory
cytokines such as IL-6 and TNF-
The signaling pathway predicted by network pharmacology was further validated
using ELISA assay and WB analysis. Our results demonstrate M. delavayi
restores the function of the NO–sGC–PKG signaling pathway by elevating nNOS activity
to recover NO levels, thereby increasing the levels of sGC and PKG to exert
antioxidant effects through the scavenging of free radicals and inhibition of
lipid peroxidation. Oxidative stress can oxidize the heme group of sGC from the
ferrous to the ferric state or induce heme dissociation, markedly impairing sGC
responsiveness to NO. Such disruption compromises smooth muscle relaxation,
anti-inflammatory responses, and mucosal barrier integrity [37, 38]. M.
delavayi may recover sGC activity through multiple mechanisms, including free
radical scavenging, enhancement of NOS-mediated NO production, and reversal of
sGC heme oxidation [39, 40, 41]. Then, the effect of M. delavayi on sGC could
elevate cGMP level and reactivate PKG and downstream effectors. Unexpectedly,
high-dose M. delavayi markedly increased iNOS expression, which may
reflect a compensatory or context-dependent regulatory response rather than a
purely pathogenic effect. Several mechanisms, including compensatory feedback to
altered NO–ROS coupling or activation of transcriptional pathways known to
regulate iNOS expression, may contribute to this response. However, these remain
speculative and require further experimental verification. Importantly,
M. delavayi simultaneously reduced O₂⁻ levels, thereby limiting the
formation of ONOO⁻ and diverting NO from pathological nitrative reactions toward
physiological NO–sGC–PKG signaling pathway. Collectively, these effects contribute to
cytoprotective outcomes. Moreover, previous reports indicate that polysaccharides
derived from M. delavayi roots can activate NF-
This study has several limitations. First, the findings are based on an L-arginine-induced mouse model, which may not fully represent the complexity of human functional dyspepsia. Second, although multiple targets and pathways were predicted by network pharmacology, only the NO–sGC–PKG signaling pathway was experimentally validated. Finally, the specific active compounds of M. delavayi responsible for the observed effects were not identified. Future studies, including clinical validation and component-level analysis, are needed to confirm these findings.
In summary, M. delavayi effectively ameliorates L-arg-induced gastrointestinal dysfunction through a multifaceted mechanism involving the regulation of gastrointestinal hormone secretion, restoration of the NO-sGC-PKG signaling pathway, and maintenance of redox homeostasis.
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
LS and BJ designed the research study. LC, ZD, HW and JZ performed the research. BJ and LS provided guidance on experimental procedures. JW, LC and ZD conducted data analysis and visualization. LC, and ZD developed the methodology. LS and BJ acquired funding. LC and ZD drafted the original manuscript. JW, HW and JZ conducted literature search and reference collection. LC and ZD reviewed and edited the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All animal experiments were conducted in accordance with the approved protocols and guidelines of the Animal Ethics Committee of Dali University (Ethics No. 2024-P2-110). All animal euthanasia procedures were performed in accordance with the AVMA Guidelines for the Euthanasia of Animals.
We extend our heartfelt gratitude to all participants, researchers, technicians, and reviewers who contributed to this study. Their invaluable support and contributions have been instrumental in the successful completion of this research.
This research received no external funding.
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/IJP48017.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.