











 , Yujie Wang 2, Chongjun Zhao 2, Xinfang Xu 2, Xiangri Li 2,*
, Yujie Wang 2, Chongjun Zhao 2, Xinfang Xu 2, Xiangri Li 2,*1 School of Pharmacy, Jiangxi University of Chinese Medicine, 330004 Nanchang, Jiangxi, China
2 Research Center of Traditional Chinese Medicine Processing/Beijing Key Laboratory for Quality Evaluation of Chinese Medicine, Beijing University of Chinese Medicine, 102400 Beijing, China
Abstract
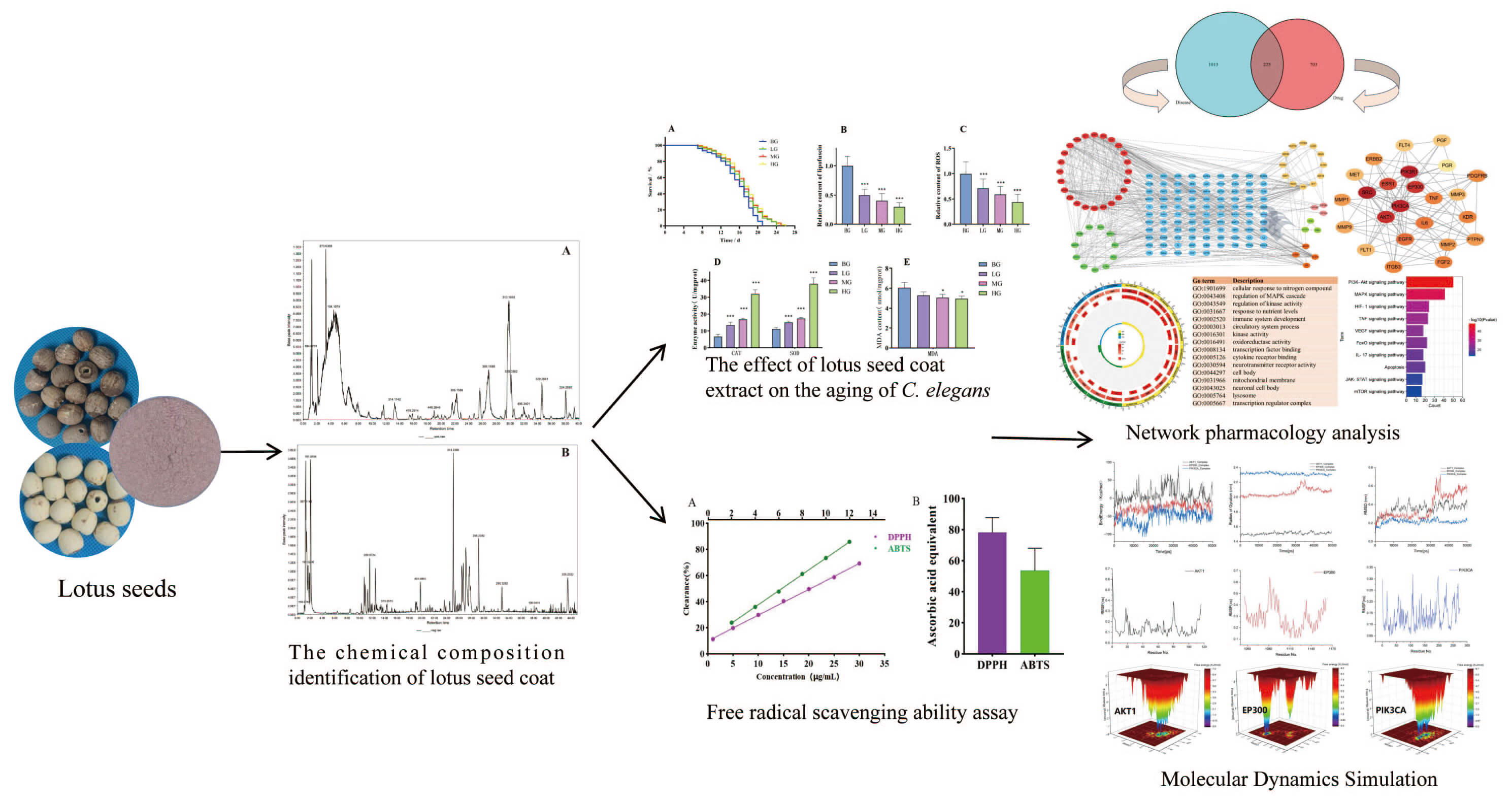
Aging is a critical global public health challenge, and natural products represent a key area in anti-aging research. Therefore, this study aimed to explore the chemical composition and anti-aging efficacy of the lotus seed coat.
Ultra-high-performance liquid chromatography-Orbitrap mass spectrometry (UHPLC/Orbitrap-MS) was used to analyze chemical components. The lifespan of Caenorhabditis elegans (C. elegans) treated with lotus seed coat extract was measured, along with oxidative stress markers and antioxidant enzyme activity. Network pharmacology and molecular dynamics simulations were employed to predict any associated anti-aging mechanisms.
A total of 67 compounds were tentatively identified in the lotus seed coat, including 14 alkaloids, 19 flavonoids, and 6 phenolic acids. Lotus seed coat extract prolonged the lifespan of C. elegans (p < 0.05), reduced lipofuscin and reactive oxygen species (ROS) levels (p < 0.001), increased superoxide dismutase (SOD) and catalase (CAT) activity, and decreased malondialdehyde (MDA) content. Network pharmacology linked the anti-aging effects of the lotus seed coat extract to the regulation of SRC, PIK3R1, AKT1, IL6, and TNF genes expression and implicated the phosphatidylinositol 3-kinase/protein kinase B pathway (PI3K–Akt), mitogen-activated protein kinase (MAPK), hypoxia-inducible factor 1 (HIF-1), forkhead box O (FOXO), tumor necrosis factor (TNF), and mammalian target of rapamycin (mTOR) signaling pathways. Molecular simulations confirmed that neferine strongly binds to phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit alpha (PIK3CA), histone acetyltransferase p300 (EP300), and RAC-alpha serine/threonine-protein kinase (AKT1).
Lotus seed coat is rich in bioactive compounds and exhibits significant anti-aging potential.
Graphical Abstract
Keywords
- lotus seed coat
- anti-aging
- Caenorhabditis elegans
- antioxidant
- network pharmacology
Aging is characterized by a gradual decline in bodily functions, leading to increased susceptibility to cardiovascular and neurodegenerative diseases, which can have profound impacts on the overall health of elderly individuals [1, 2]. Research on anti-aging has been ongoing throughout history, with various theories proposed to explain the mechanisms of aging. These include theories on free radicals, oxidative stress, immune aging, DNA damage, mitochondrial damage, and cell autophagy [3, 4]. While existing anti-aging agents have demonstrated certain efficacy with specific targets, they face challenges such as the limited efficacy of single-component agents, risks of side effects with long-term use, and low bioavailability [5]. Moreover, they are unable to cover the multi-dimensional mechanisms of aging. Against this backdrop, the exploration of highly effective, low-toxicity, and multi-target synergistic anti-aging drugs from natural products has emerged as a key research trend that also aligns with the health needs of “holistic intervention and long-term safety [6, 7]”.
Lotus seeds are dried, fully matured seeds from the aquatic plant Nelumbo nucifera Gaertn. They are widely acknowledged as having purported longevity-enhancing bioactivities [8, 9]. Lotus seed coat is a by-product of lotus seed processing, accounting for approximately 15% of the total weight of lotus seeds [10]. It is typically discarded as waste and casually disposed of in ditches or along roadsides. The lotus seed coat corresponds to the traditional medicinal term Lianyi [11]. It consists of the membranous structure attached to the surface of the lotus seed kernel (endosperm) after the outer hard shell of the lotus seed has been removed. It appears reddish-brown when dried [12, 13]. According to Bencao Qiuyuan, lotus seeds are described as “tonifying when peeled; securing essence and invigorating the spleen when used with the coat”. This suggests the lotus seed coat can prevent the dissipation of qi, blood, and body fluids, thereby exerting an anti-aging effect [14]. Research on the lotus seed coat has gradually increased in recent years, particularly with regard to its chemical composition and antioxidant capabilities. However, the potential association between the lotus seed coat and longevity still needs further investigation.
Driven by the emerging interest in the anti-aging potential of lotus seed coat, we designed a multi-layered study to systematically investigate its bioactive components, antioxidant capacity, and in vivo anti-aging effects, followed by analysis of the predicted mechanism. First, we employed HPLC/Orbitrap-MS to characterize the chemical composition of the lotus seed coat. This step was critical for linking specific compounds to subsequent biological effects. Since excessive accumulation of free radicals is a well-documented driver of cellular senescence and aging, we then evaluated the in vitro antioxidant capacity of the extract using 2,2-diphenyl-1-picrylhydrazyl (DPPH) and 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) assays. The results of these assays provided preliminary validation of its anti-aging potential.
To further confirm the effects in a living system, C. elegans was selected as an in vivo model. C. elegans is a widely accepted and utilized organism for anti-aging research due to its short lifespan, genetic tractability, and conservation of key aging-related pathways. Core aging-related endpoints were evaluated following exposure of wild-type N2 worms to graded concentrations of lotus seed coat extract. These included organism lifespan, the levels of lipofuscin (a senescence biomarker) and reactive oxygen species (ROS), the activity of antioxidant enzymes (superoxide dismutase [SOD], catalase [CAT]), and the level of malondialdehyde (MDA), a lipid peroxidation marker. Finally, to link the observed phenotypic effects with molecular mechanisms, network pharmacology analysis was performed to predict potential targets and signaling pathways underlying the anti-aging activity of lotus seed coat. This computational approach identified key molecular mediators, while also providing a theoretical foundation for future mechanistic validation studies.
The reference standard, including neferine, quercetin, and kaempferol, was dissolved in methanol at a concentration of 0.1 mg/mL. All solutions were stored at –20 °C until use.
Professor Xiangri Li, an expert in Chinese materia medica from Beijing University of Chinese Medicine, identified the lotus seeds as the dried mature nuts of Nelumbo nucifera Gaertn., a plant belonging to the genus Nelumbo (family Nymphaeaceae). The lotus seed coat (Fig. 1) has been deposited in the Laboratory of Chinese Materia Medica Processing, Beijing University of Chinese Medicine, and is available for inspection and verification by peers.
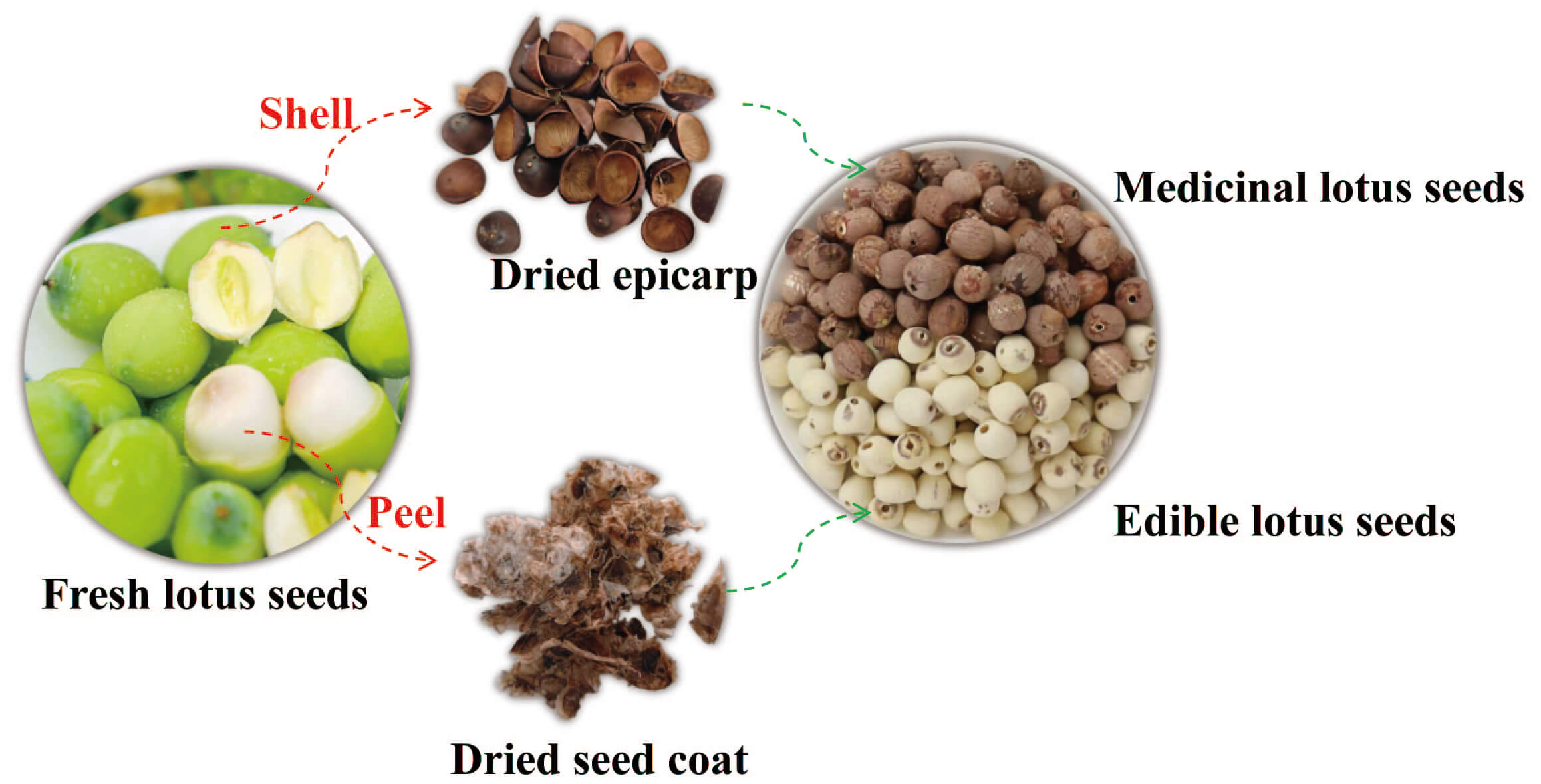 Fig. 1.
Fig. 1.
Processing pathways and morphological differences: Medicinal vs. Edible Lotus Seeds.
For positive ion mode analysis, 1.0 g of the powder was extracted with 0.3
mol
A Vanquish UHPLC system (Thermo Fisher Scientific, San Jose, CA, USA) was
employed for separation. Samples were separated on an Acquity HSS T3 column (2.1
Data were processed with Xcalibur TM 4.1 software (Thermo Fisher). MS/MS data were further analyzed using an online LC/MS database (http://www.hmdb.ca; https://massbank.eu/MassBank/), and by comparing the MS/MS spectra with reference standards and the literature.
One gram of lotus seed coat powder was weighed and extracted with 20 mL of 70% methanol (v/v) for 60 min in an ultrasonic water bath (40 kHz, 500 W). The extracts were passed through filter paper and stored at 4 °C. The extraction rate of the lotus seed coat is approximately 22.4%.
Free radicals (atoms or groups with unpaired electrons) are closely related to aging. Common methods for measuring free radical scavenging capacity include the DPPH and ABTS assays. The DPPH solution employed in the assay was prepared at a concentration of 0.4 mmol/L, with 70% methanol serving as the diluent. For the standard group, 100 µL of ascorbic acid standard solution was added to 100 µL DPPH solution. For the sample group, 100 µL of lotus seed coat sample extract diluent was added to 100 µL of DPPH solution. The reaction system was maintained at room temperature under dark conditions for 30 minutes, after which the absorbance was measured at 517 nm using a microplate reader.
The ABTS
Aχ: absorbance of the sample solution/standard solution after the reaction.
A0: absorbance of the blank reaction.
The DPPH/ABTS clearance of the sample was calculated by fitting the equation to the standard curve, and then expressed as ascorbic acid equivalent: mg ascorbic acid eqg DW (mgAAE/g DW).
The cultivation and experimental manipulation of C. elegans were
conducted in accordance with established protocols [15]. Freeze-dried powder of
70% methanol extract from the coat of lotus seeds was dissolved in water and
added to the nematode growth medium (NGM) plate. Simultaneously, E. coli
OP50 was spread on the NGM plate to serve as nourishment for C. elegans.
In preliminary toxicological experiments, the lotus seed coat was found to have
no toxic effect on C. elegans in the concentration range of
0.01~1 mg
C. elegans at the L4 stage were chosen and transferred onto NGM plates (3 plates per group, 30 strips per plate). To maintain a consistent drug concentration, C. elegans were transferred to new NGM plates the following day, and the number of deceased individuals was recorded. The blank group was used as the control, and survival time was calculated from the transfer time until all C. elegans had died, which marked the end of the experiment. The experiment was repeated three times. The lifespan of each group of C. elegans was recorded and used to calculate the average, median, and maximum lifespans. Evaluation criteria for the death of C. elegans were no movement or swallowing action, and no response after light touch.
All experiments used synchronized L4-stage C. elegans under uniform conditions. For the lipofuscin assay, worms were transferred to fresh NGM plates (3 plates/group, 30 worms/plate) and cultured for 5 days, then rinsed three times with M9 buffer, anesthetized, and mounted on slides. Blue fluorescence intensity was then recorded via an inverted microscope and analyzed with ImageJ 1.54f (National Institutes of Health, Bethesda, MD, USA). For the ROS assay, worms were cultured as above, then rinsed, centrifuged (supernatant discarded), and incubated with 200 µL of 100 µM DCFH-DA (Solarbio, ID31309, Beijing, China) for 120 min. Green fluorescence intensity was then recorded via an inverted microscope and analyzed with ImageJ. For the SOD/CAT activity and MDA content assay, worms were transferred to drug-supplemented NGM plates and cultured for 5 days, then rinsed, centrifuged, repeatedly washed with M9 buffer, and stored at –80 °C. For assessment, the samples were ground, centrifuged, and the supernatant analyzed using kits (A001-3-2, A003-1-2, A007-2-1, Nanjing Jiancheng, Nanjing, China) as per the manufacturer’s instructions.
Targets for the lotus seed coat compounds were predicted using two databases: SEA (https://sea.bkslab.org/) and SwissTargetPrediction (http://www.swisstargetprediction.ch/). The chemical composition of the lotus seed coat was determined based on in-house experimental results [10]. Candidate targets were standardized to UniProtKB IDs and corresponding gene symbols via the UniProtKB database. Aging-related targets were retrieved from differential genes identified using the Genecards database (https://www.genecards.org/). Common targets were defined as the intersection of compound-predicted targets and aging-related targets [16].
To construct the PPI network, UniProt IDs of common targets were uploaded to the STRING database. The species was set as Homo sapiens, and the interaction confidence score threshold was adjusted to 0.90 (highest confidence). The resulting PPI network was visualized using Cytoscape 3.9.1 (Cytoscape Consortium, Seattle, WA, USA), followed by topological analysis via the CytoHubba and MCODE plug-ins to identify key nodes and functional modules.
Metascape (http://metascape.org/) is a robust platform enabling users to apply currently popular bioinformatics analysis methods to bulk gene and protein analysis for knowledge of gene or protein function. The gene names for the above vital targets were entered into the Metascape website, and species were limited to humans. This enabled Gene Ontology (GO) functional annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis using the Metascape website.
Molecular docking was performed using Autodock Vina 1.2.5 (The Scripps Research Institute, La Jolla, CA, USA) to assess the binding affinity of small molecules to the key protein. Subsequently, three representative small molecule-protein complexes were selected for molecular dynamics (MD) simulations to evaluate complex stability. The main procedures were carried out as follows:
① The AMBER14 force field was employed to optimize the energy of the
protein-ligand system. A periodic boundary simulation box was constructed
(expanded by 10 Å), filled with TIP3P water molecules (density: 0.997
g
② The simulation was conducted over a duration of 50,000 ps (50 ns) at a temperature of 310 K (NVT ensemble with Langevin temperature control) to simulate physiological conditions, with a pH of 7.4 (physiological condition). The initial structure was optimized using the default steepest descent method (500 steps).
③ Key output parameters included: Root Mean Square Deviation (RMSD) to assess overall conformational stability; Root Mean Square Fluctuation (RMSF) to analyze local dynamic fluctuations of residues in the active site; Radius of Gyration (Rg) to measure protein compactness; binding energy to evaluate the energy released upon ligand-protein binding; and Free Energy Landscape (FEL, also referred to as free energy surface) to investigate processes such as protein folding and molecular motion, thus helping to understand molecular stability and dynamic behavior across different states.
Standard curves were analyzed using linear regression. All statistical analyses
were performed using GraphPad Prism 8.3.0 (Boston, MA, USA). The ANOVA method was
used for statistical comparisons between groups. All data are the mean of three
replicate determinations, presented as the mean
The chemical constituents of the lotus seed coat were analyzed by UHPLC/Orbitrap-MS, with the base peak chromatogram shown in Fig. 2. Data interpretation was performed using online LC/MS databases (HMDB: https://www.hmdb.ca/; MassBank: https://massbank.eu/MassBank/) and by comparison with literature-reported compounds. A total of 67 compounds were tentatively identified, comprising 14 alkaloids, 19 flavonoids, 6 phenolic acids, 12 fatty acids, 2 lactones, 7 saccharides, and 7 tannins. Among these, quercetin, kaempferol, and neferine were confirmed by comparison with authentic standards. Detailed information is provided in Supplementary Tables 1,2.
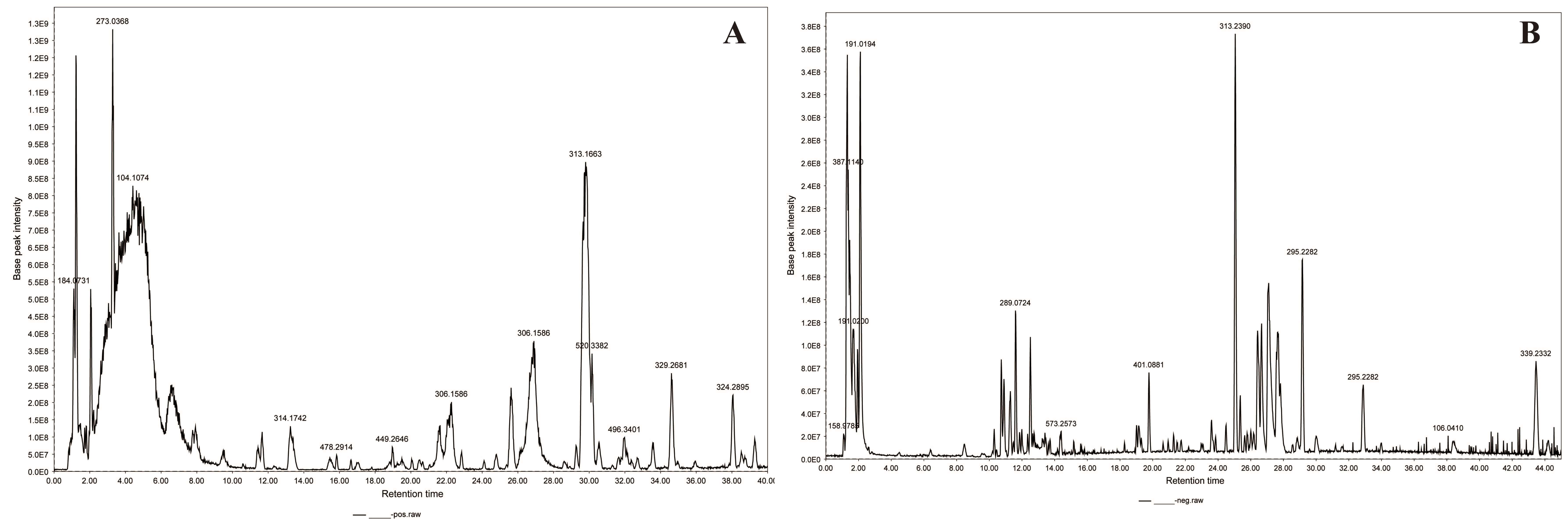 Fig. 2.
Fig. 2.
Base peak chromatograms of lotus seed coat. (A) positive ion modes; (B) negative ion modes
The inferred compounds are shown in Supplementary Materials and were
derived from references and online databases [11, 12, 17]. The main alkaloid
components in lotus seed coat are classified into aporphine-type,
mono-benzylisoquinoline-type, and bis-benzylisoquinoline-type alkaloids. Cleavage
of bis-benzylisoquinoline alkaloids primarily involves the fragmentation of two
benzyl groups [12, 18]. Typically,
In general, flavonoid glycosides, with the exception of carbon glycosides, undergo initial cleavage at the glycosidic bond to generate fragment ions of flavone aglycones that have lost their glycosyl groups [20]. Subsequently, these fragment ions undergo cleavage like that of flavonoids. The most prominent cleavage pathway for flavonoids is Retro-Diels-Alder (RDA) cleavage of the C ring, with positions 1 and 3 of the C ring being commonly cleaved to generate an ion containing the m/z 151 of the A ring. Upon comparison with a reference standard, compounds 38 and 41 (Supplementary Table 2) were identified as quercetin and kaempferol [21].
Proanthocyanidins are a class of polymers consisting of flavan-3-ols condensed through C-C bonds. They typically have catechins or epicatechins as the basic unit, with the primary mode of cleavage being rupture of the covalent bonds between the constituent units [22, 23]. Like flavonoids, the primary mode of cleavage of flavan-3-ols is also RDA cleavage of the C ring. The molecular ion peak is m/z 577.1351, and secondary fragment ions include m/z 425.0881, m/z 407.0772, m/z 289.0718, and m/z 161.0243.
Most phenolic acids contain carboxyl or phenolic hydroxyl groups, readily losing neutral molecules such as H2O, COOH, and CO2 under high-energy collisions [24]. Ferulic and caffeic acids readily lose acyl groups. In the case of ferulic acid, for example, the molecular ion peak is m/z 193.0504 in negative ion mode, and the secondary fragment ions are m/z 178.0272, m/z 149.0608, and m/z 134.0373.
The fatty acids contained in the lotus seed coat are mostly straight-chain unsaturated fatty acids. Hydroxy-octadecenoic acid was the most abundant among the fatty acids identified in this experiment. The cleavage pathway of the fatty acids is generally straightforward, with easy loss of H2O or CO fragments, and firm molecular ionic peaks [11].
Linear regression analysis was performed using clearance as the vertical coordinate and the ascorbic acid control mass concentration (µg/mL) as the horizontal coordinate (Fig. 3). In the DPPH method, ascorbic acid was linear in the range 1~30 µg/mL, y = 0.0198x + 0.0983, R2 = 0.9995. Similarly, in the ABTS method, ascorbic acid was linear in the range 2~12 µg/mL, y = 0.0624x + 0.0802, R2 = 0.9997 (Fig. 3A).
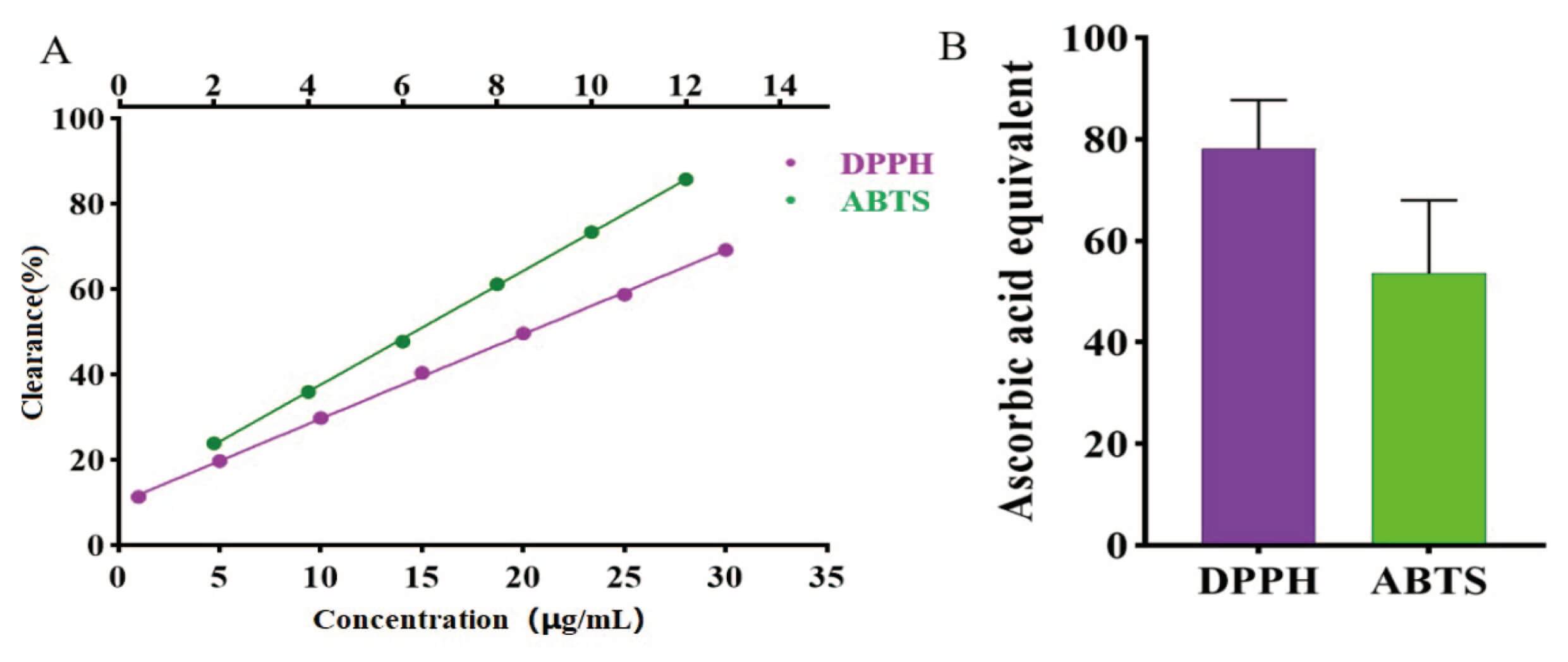 Fig. 3.
Fig. 3.
Assay results for in vitro free radical scavenging ability of the lotus seed coat. (A) Ascorbic acid standard curve. (B) Statistical analysis of the ascorbic acid equivalent content of the lotus seed coat. DPPH, 2,2-diphenyl-1-picrylhydrazyl; ABTS, 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid).
A 70% methanolic extract of lotus seed coat exhibited potent in vitro free radical scavenging ability, expressed as ascorbic acid equivalents. With the DPPH and the ABTS methods, the ascorbic acid equivalents of lotus seed bark were 78.25 mg/g and 53.59 mg/g, respectively (Fig. 3B). The results showed that the lotus seed coat had good free radical scavenging ability in vitro, indicating that it also had good anti-aging activity [25]. Due to the limitations of in vitro experiments, we also carried out in vivo anti-aging experiments.
Based on experimental data, we calculated the average, median, and maximum
lifespan of C. elegans. The C. elegans lifespan curve is shown
in Fig. 4A. Compared with the blank group, various doses of lotus seed coat
extract were found to increase the average lifespan of C. elegans. The
average lifespan in the blank group was 14.31 days, compared to 15.11, 15.97, and
15.82 days in the low-, medium-, and high-dose groups of lotus seed coat extract,
respectively (Fig. 4A). The results were significantly different
(p
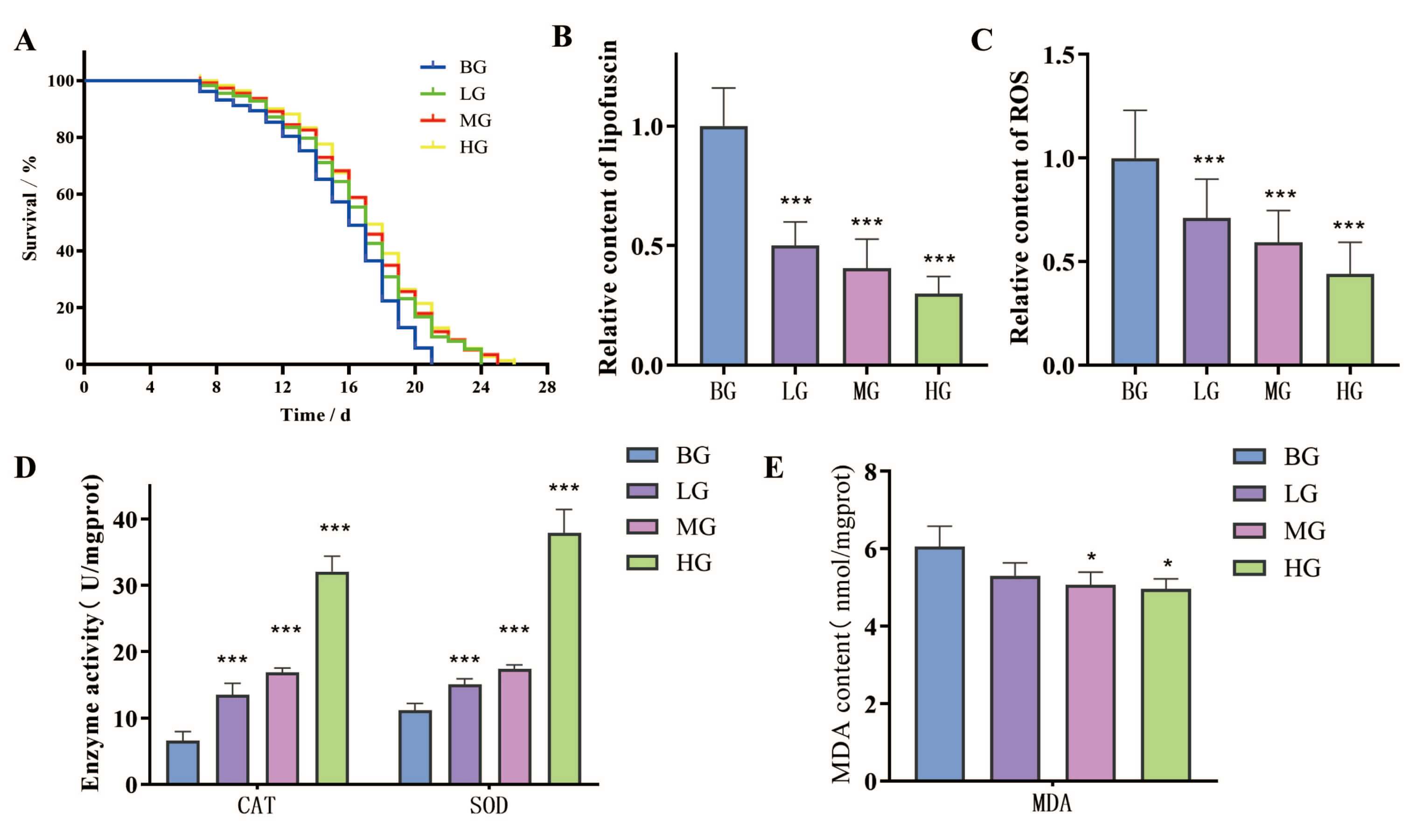 Fig. 4.
Fig. 4.
The effect of lotus seed coat extract on the aging of
C. elegans. (A–E) The effects of lotus seed coat extract on the
lifespan, lipofuscin content, reactive oxygen species (ROS) levels, antioxidant
enzyme (SOD/CAT) activities, and Malondialdehyde (MDA) content in C.
elegans. (*p
The lipofuscin content of C. elegans accumulates with age, with its
autofluorescence properties used as a sign of aging [26]. Lotus seed coat extract
significantly reduced the lipofuscin level in the nematode body (Fig. 4B and Fig. 5). The level was 49.99%, 59.58% and 70.12% lower in the low-, medium-, and
high-dose groups compared to the blank group, respectively (p
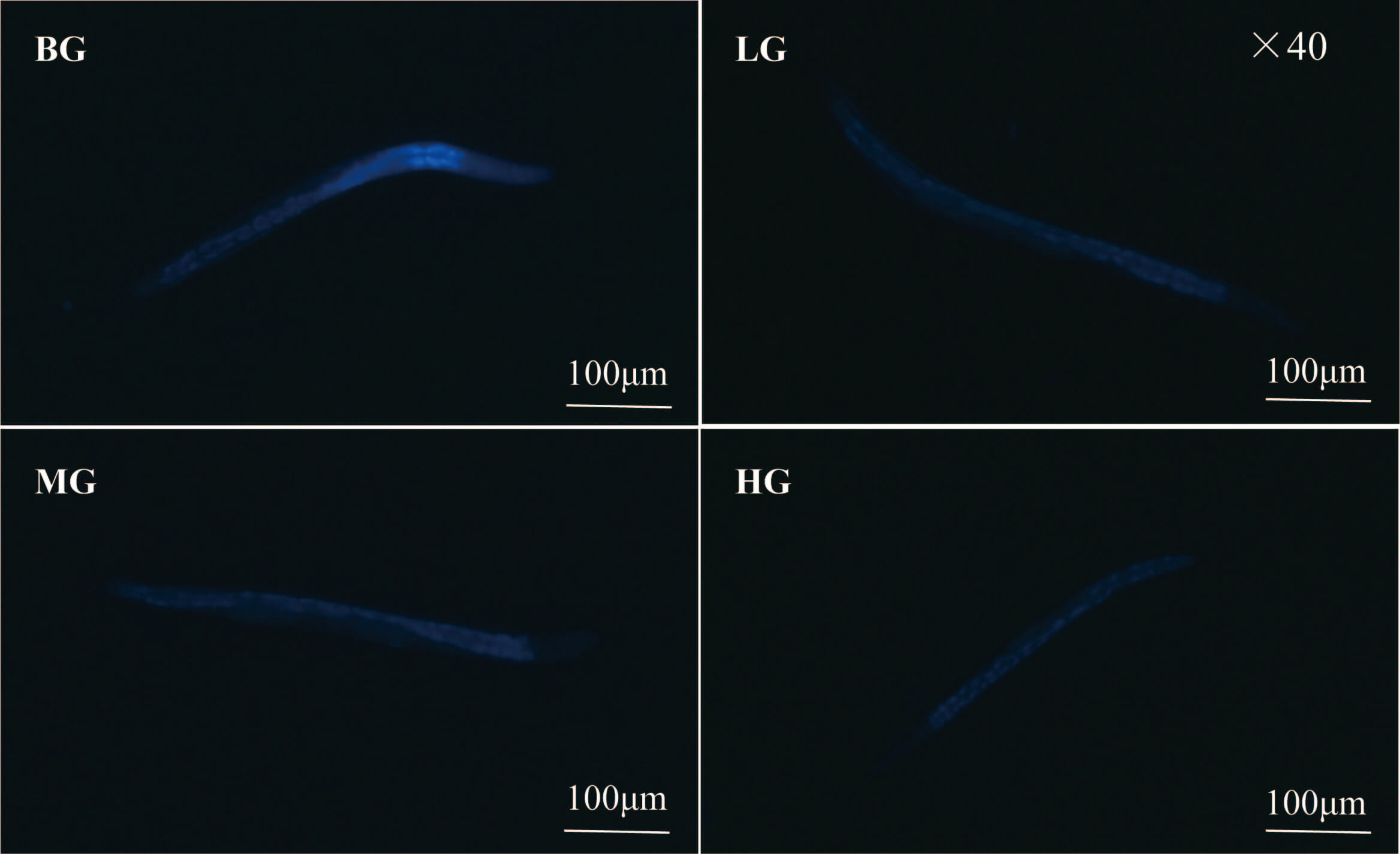 Fig. 5.
Fig. 5.
Lipofuscin fluorescence image of C. elegans. Scale bar = 100 µm.
The ROS content in the body of C. elegans increases with the aging
process. Excessive accumulation of ROS in the body can activate aging-related
signaling pathways, inducing cell cycle arrest and the acquisition of a
senescence-associated secretory phenotype (SASP) in cells, thereby accelerating
the aging process [27]. Lotus seed coat extract significantly reduced the ROS
level in C. elegans, as shown in Fig. 4C and Fig. 6. Lipofuscin levels
in C. elegans were reduced by 28.92%, 40.87%, and 56.12% in the low-,
medium-, and high-dose groups, respectively, compared to the blank group
(p
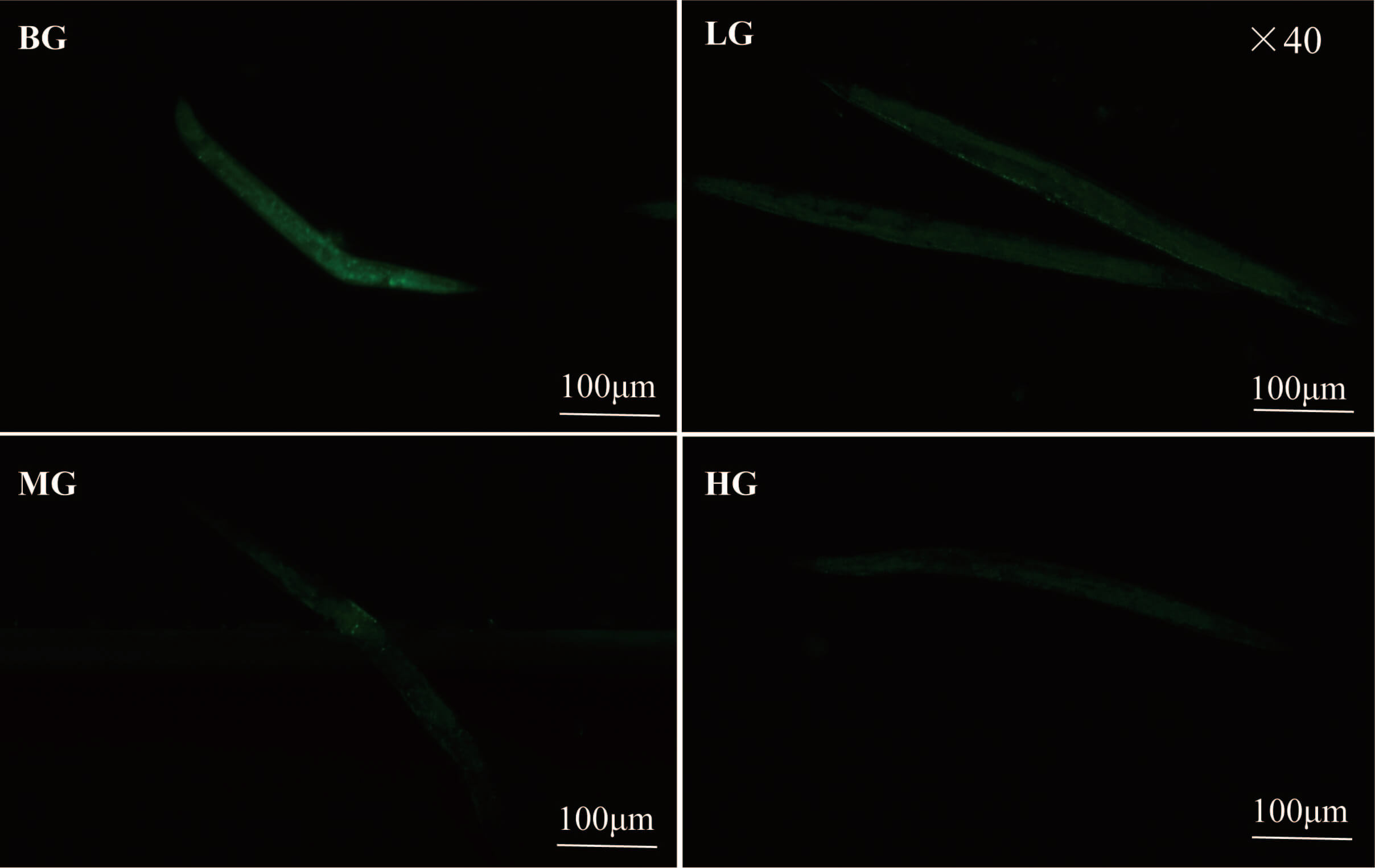 Fig. 6.
Fig. 6.
ROS fluorescence image of C. elegans. Scale bar = 100 µm.
SOD and CAT are the main antioxidant enzymes in C. elegans and can
eliminate and reduce excessive free radicals, thereby delaying aging. MDA is one
of the essential biomarkers of the oxidative stress response and can also reflect
the aging status [28]. The results for SOD and CAT activity in the C.
elegans body are shown in Fig. 4D. Lotus seed coat extract
significantly improved the antioxidant enzyme activity in the C. elegans
body in a dose-dependent manner (p
After screening and removing duplicate targets, the 67 compounds identified by
mass spectrometry in lotus seed coats were predicted to have 672 drug targets
(Fig. 7). Disease targets were retrieved from Genecard databases using the
keyword “aging”. Those with a relevance score
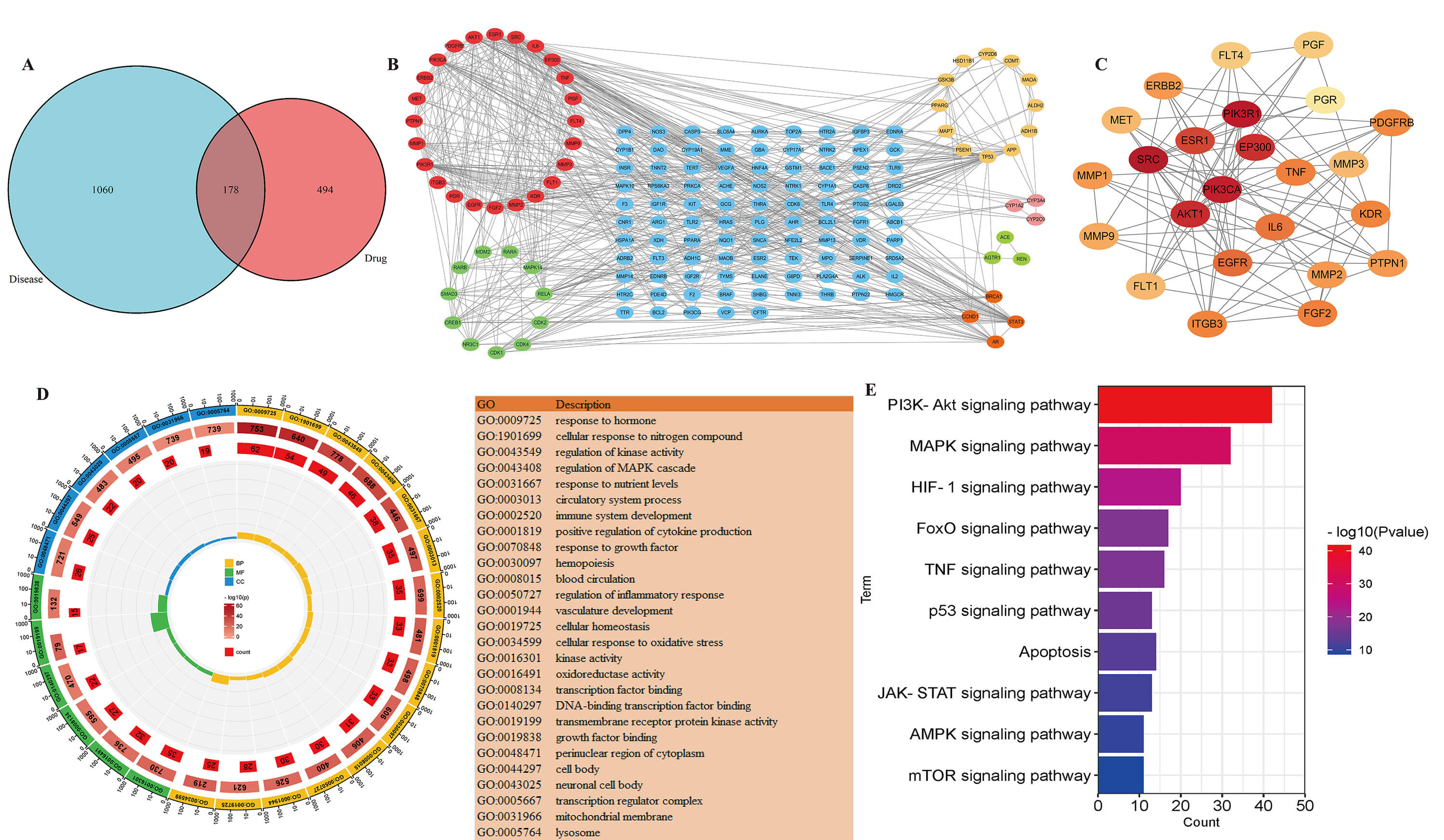 Fig. 7.
Fig. 7.
Results of network pharmacology analysis. (A) Common targets of the compound and of aging. (B) A protein-protein interaction network with a common target. Different colors represent different clustering modules created by the MCODE algorithm. (C) In the first module, the hub targets were grouped. (D) GO enrichment analysis of common targets. (E) KEGG enrichment analysis of common targets. GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes.
Common targets were first imported into the STRING database. Those without
interactions were removed, and high-confidence targets with a threshold value
In the PPI network, the degree of freedom for each node represents the number of interactions with other nodes. The higher the value for the degree of freedom, the higher the importance of that node. The first 10 core genes predicted by this analysis were SRC, PIK3R1, PIK3CA, AKT1, EP300, ESR1, EGFR, IL6, TNF, and FGF2.
GO and KEGG pathway enrichment analyses were performed to analyze the biological functions and pathways involved in the 225 common targets. GO functional annotation enrichment analysis focused on three aspects: biological processes (BP), cellular composition (CC), and molecular functions (MF). The analysis identified 2016 BP, including regulation of hormone levels, regulation of nitrogenous compounds in cells, regulation of enzyme activity, and regulation of the MAPK cascade reaction; 122 CC including the cytoplasmic perinuclear region, cell bodies, transcriptional regulator complexes, mitochondrial membranes, and lysosomes; and 194 MF including oxidoreductase activity, transcription factors, protein kinase activity, and growth factors (Fig. 7D). GO functional annotation enrichment analysis revealed that the anti-aging effect of lotus seed coat is mediated mainly through biological processes related to hormone levels, enzyme activity, oxidative stress, immunity, inflammation, and blood.
KEGG pathway enrichment analysis indicated the anti-aging effect of lotus seed coat was mainly related to autophagy and to the phosphatidylinositol 3-kinase/protein kinase B pathway (PI3K–Akt), mitogen-activated protein kinase (MAPK), hypoxia-inducible factor 1 (HIF-1), forkhead box O (FOXO), tumor necrosis factor (TNF), and mammalian target of rapamycin (mTOR) ignaling pathways (Fig. 7E).
Neferine was identified as the primary alkaloid component in the lotus seed coat
[29]. Analysis by high-performance liquid chromatography (HPLC) revealed that its
content in the lotus seed coat was 0.033 mg/g (Supplementary Fig. 1). We
next selected the top six key proteins in terms of degree value for molecular
docking analysis with neferine. The binding energy of each protein to neferine
was found to be
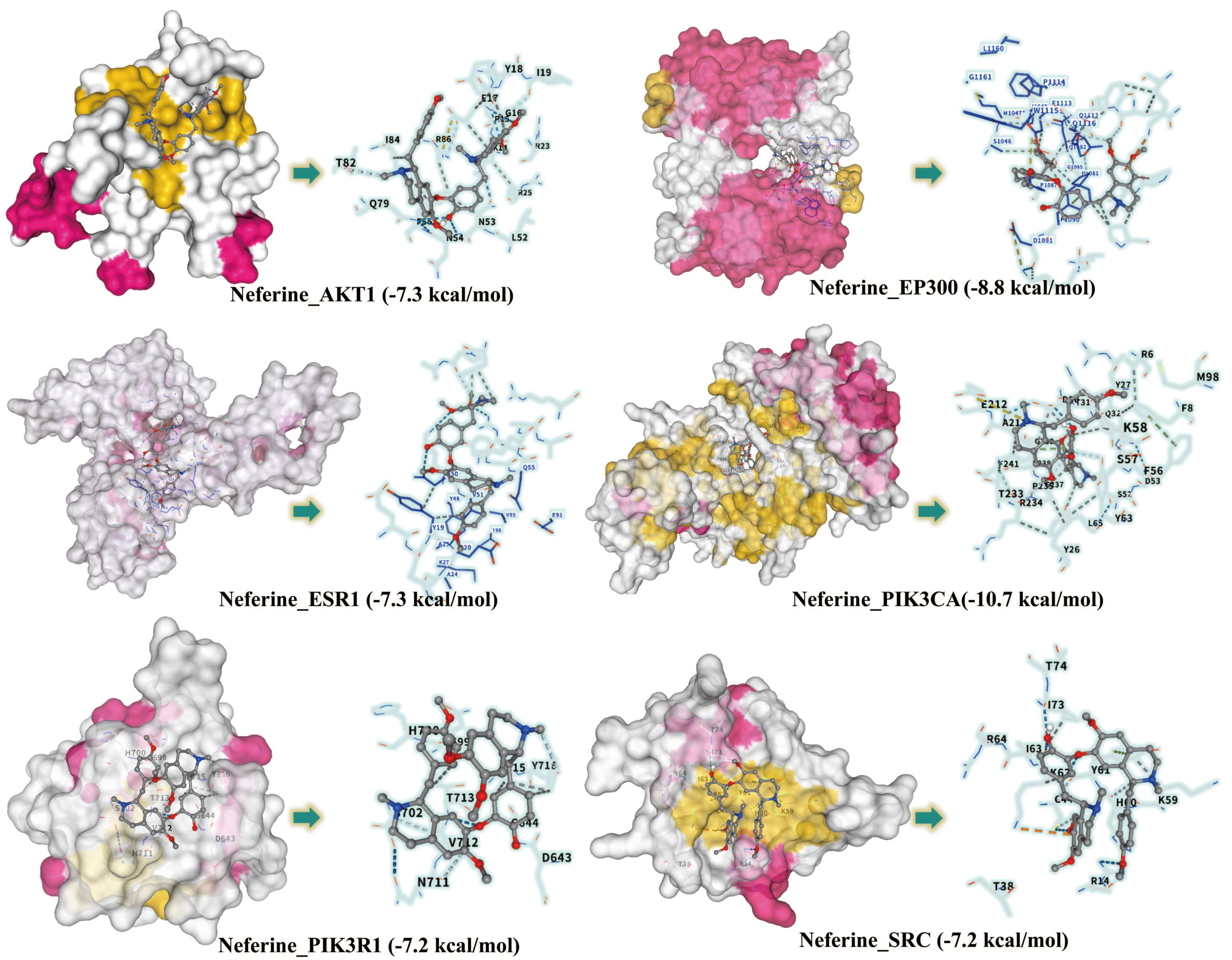 Fig. 8.
Fig. 8.
Binding modes and affinities of Neferine with six core proteins.
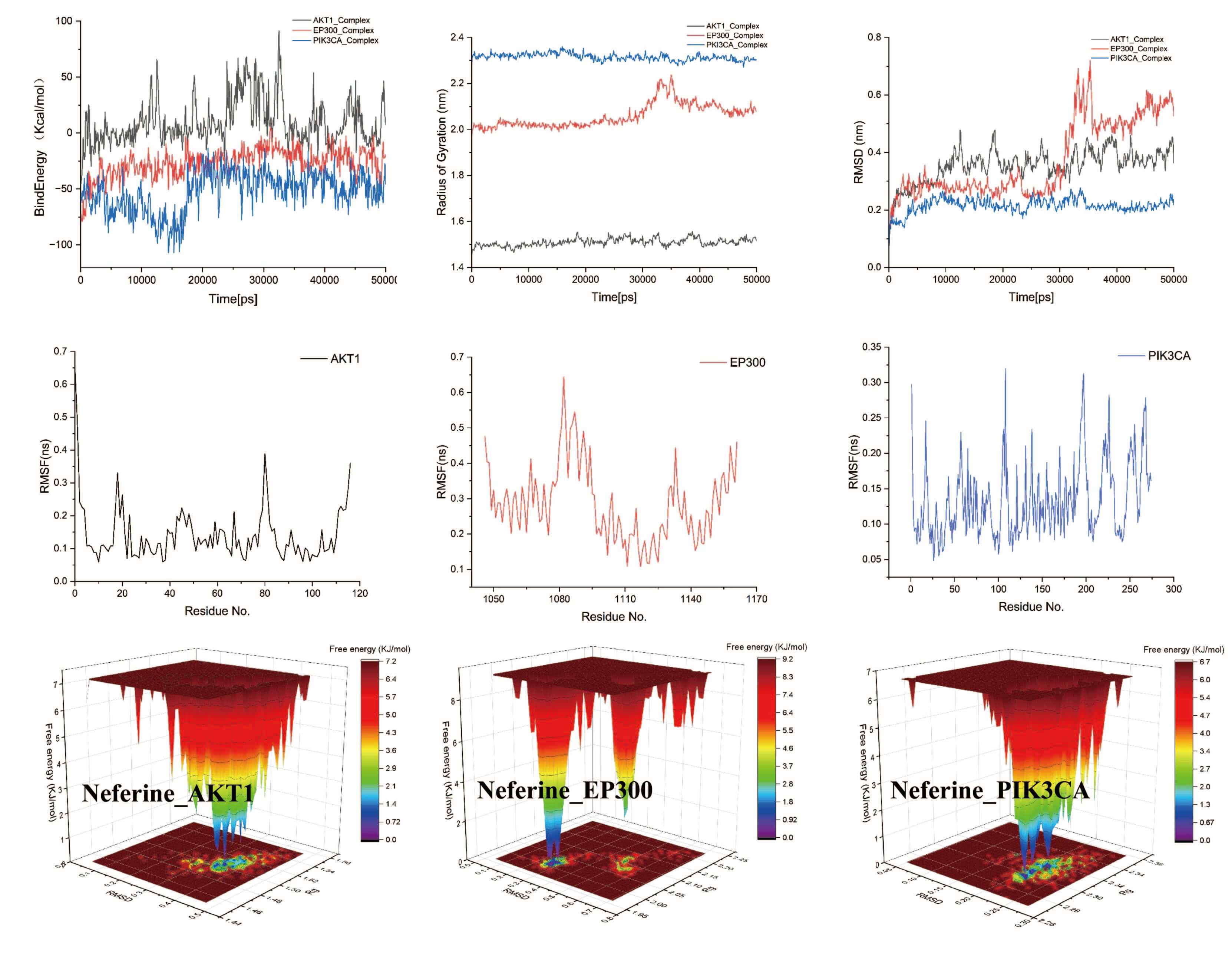 Fig. 9.
Fig. 9.
Molecular dynamics simulation results for Neferine with PIK3CA, EP300, and AKT. PIK3CA, phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit alpha; EP300, histone acetyltransferase p300; AKT1, RAC-alpha serine/threonine-protein kinase.
Aging is a natural physiological process in organisms characterized by the gradual accumulation of cellular damage and metabolic waste, leading to functional decline, increased vulnerability, and ultimately organismal death [30]. To date, significant progress has been made in anti-aging therapeutics, particularly in the development of natural and synthetic compounds. These anti-aging strategies and compounds are primarily focused on the clearance of senescent cells, augmentation of NAD, anti-inflammatory and antioxidant defense, restoration of proteostasis, and activation of telomerase [31].
In the current study, 67 components of the lotus seed coat were inferred by LC-MS, including 14 alkaloids, 19 flavonoids, 6 phenolic acids, 12 fatty acids, and several other compounds. Our results showed that the lotus seed coat can eliminate free radicals in vitro, indicating that it may have robust anti-aging effects. Lotus seed coat extract was found to significantly prolong the lifespan of the experimental model C. elegans. Moreover, it reduced the levels of lipofuscin and ROS, increased the activities of SOD and CAT, and reduced the level of MDA in this model. In summary, treatment with lotus seed coat extract resulted in moderate but significant anti-aging-related phenotypes in C. elegans, possibly by attenuating oxidative stress regulatory pathways.
To link the observed phenotypic effects of lotus seed coat extract with potential molecular mechanisms, network pharmacology was employed to integrate compound-target-disease interactions. This multi-target prediction approach found that alkaloids and flavonoids may be the primary anti-aging components in the lotus seed coat. These components exert their effects by regulating genes such as SRC, PIK3R1, AKT1, IL6, and TNF through the PI3K–Akt, MAPK, and mTOR signaling pathways. Previous studies have shown these pathways act as core regulators of the aging process by modulating oxidative stress, inflammatory responses, metabolic homeostasis, and cell survival [32, 33]. Natural products and common anti-aging agents can inhibit overactivation of the pathways through multi-target effects, thereby providing potential strategies for delaying aging [34].
As a natural product, lotus seed coat extract has the advantages of low effective dose, minimal side effects, and stable efficacy, making it suitable for long-term intervention [35]. Its core mechanisms may overlap with existing anti-aging products, including antioxidant activity and the regulation of classical pathways such as PI3K–Akt/mTOR and FOXO. Lotus seed coat may delay aging by scavenging free radicals, activating endogenous antioxidant systems, and modulating cellular metabolism. Notably, its uniqueness lies in multi-component synergy and multi-target network regulation, forming a composite mode of “holistic intervention-precise regulation” that is particularly suited for natural, long-term aging prevention. This addresses the limitations of single-component anti-aging products, including narrow target scope, low bioavailability, or prominent side effects [36]. Furthermore, this study found that by-products of lotus seed processing can be converted into anti-aging resources, thereby providing a practical example for the high-value utilization of agricultural waste and demonstrating both academic innovation and industrial application.
However, this study did not clarify whether the extract activates classic anti-aging pathways, nor did it validate the observed metabolic regulatory effects in mammalian models. Despite these limitations, it also provided initial assessment and prediction, thereby laying the groundwork for future research endeavors. As the focus of this study was mainly on preliminary evaluation and prediction, further experimental investigations are required.
This preliminary study infers that the lotus seed coat contains active antioxidant components, as determined by component analysis and in vitro antioxidant experiments. The anti-aging potential of the lotus seed coat was also verified using Caenorhabditis elegans and network pharmacology. Our results provide a replicable and practical example for the resource utilization of agricultural waste.
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
XRL: Conceptualization, Methodology, Supervision; XWY: Investigation, Formal analysis, Supervision, Funding acquisition; CSL: Data curation, Validation, Writing–review & editing; XFX and CJZ: Supervision, Data curation; YJW: Data curation, Validation. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
According to Chinese regulations and policies, this study does not involve human or animal subjects; therefore, ethical approval for the Caenorhabditis elegans experiment is not necessary. The lotus seeds used in this experiment were collected from Fuzhou City, Jiangxi Province, China. Professor Xiangri Li, an expert in Chinese materia medica from Beijing University of Chinese Medicine, identified the lotus seeds as the dried mature nuts of Nelumbo nucifera Gaertn., a plant belonging to the genus Nelumbo (family Nymphaeaceae).
The authors are thankful to the Jiangxi/Beijing University of Chinese Medicine for the assistance in conducting this study.
This study was supported by the Science and Technology Program Project of Administration of Traditional Chinese Medicine of Jiangxi Province (2024A0079), Doctoral Startup Fund (2024BSZR052).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/IJP46482.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.