



 , Yu Ding 2, Honglian Zhang 3, Amirhossein Mohammadi 4, Jayant Seth 5, Yasir Abu-Omar 6, Mohammad El-Diasty 6,*
, Yu Ding 2, Honglian Zhang 3, Amirhossein Mohammadi 4, Jayant Seth 5, Yasir Abu-Omar 6, Mohammad El-Diasty 6,*
1 School of Medicine, Queen’s University, Kingston, ON K7L 3N6, Canada
2 Institute of Biomedical Engineering, Faculty of Applied Science and Engineering, University of Toronto, Toronto, ON M5S 3E3, Canada
3 Faculty of Science, McGill University, Montreal, QC H3A 0G4, Canada
4 Faculty of Science, Toronto Metropolitan University, Toronto, ON M5B 2K3, Canada
5 School of Medicine, University of British Columbia, Vancouver, BC V6T 1Z4, Canada
6 Cardiac Surgery Department, University Hospitals Cleveland Medical Center, Cleveland, OH 44106, USA
Abstract
Cardiopulmonary bypass (CPB) is essential in cardiac surgery but is associated with significant postoperative inflammation. Epicardial adipose tissue (EAT), due to its close proximity to the myocardium and vasculature, may play a role in mediating these inflammatory responses. A systematic search of MEDLINE and EMBASE identified five studies that analyzed molecular changes in EAT, subcutaneous adipose tissue (SAT), and/or serum before and after CPB. Outcomes included changes in mRNA expression and protein levels of inflammatory and metabolic biomarkers. EAT demonstrated increased expression of fibroblast growth factor 21 (FGF-21), interleukin-6 (IL-6), and tumor necrosis factor-alpha (TNF-alpha) following CPB. Adipokine profiles revealed decreased EAT protein levels of leptin, adiponectin, and adipocyte fatty acid-binding protein (A-FABP), while circulating levels varied depending on patient comorbidities. Mitochondrial electron transport chain (ETC) gene expression significantly decreased in EAT but not in SAT. Endoplasmic reticulum (ER) stress markers including activating transcription factor 4 (ATF4), DNA damage inducible transcript 3 (DDIT3), activating transcription factor 6 (ATF6), and heat shock protein family A (Hsp70) member 5 (HSPA5) showed differential upregulation, particularly in patients with coronary artery disease (CAD). EAT is biologically active and contributes to both local and systemic inflammation following CPB. These biomolecular changes may underlie adverse postoperative outcomes, highlighting EAT as a potential therapeutic target to reduce CPB-associated complications.
Keywords
- biomolecules
- biomarkers
- cardiopulmonary bypass
- epicardial adipose tissue
Cardiopulmonary bypass (CPB) enables cardiac surgery on a motionless and bloodless heart by temporarily assuming the functions of the heart and lungs [1]. This is achieved by diverting venous blood to an extracorporeal circuit, where it is oxygenated and returned to the systemic circulation through cannulation sites depending on the specific cardiac operation. While CPB maintains perfusion and oxygenation, it is associated with a significant postoperative inflammatory response [1]. This response is triggered by multiple factors, such as contact activation of blood components with the artificial surface of the bypass circuit, ischemia and reperfusion injury, endotoxemia, and direct surgical trauma [2]. The ensuing inflammatory cascade involves the upregulation of pro-inflammatory cytokines and adhesion molecules, mediated by intracellular transcription factors [3]. In addition, several blood components including coagulation, fibrinolytic, complement, leukocyte, endothelial cell, and platelet systems are activated, resulting in a complex acute-phase reaction [3]. Notably, several studies have demonstrated the accumulation of different inflammatory mediators in pericardial fluid, suggesting that the epicardial adipose tissue (EAT) may play a role in the synthesis and release of these mediators [4].
EAT is a unique type of visceral fat deposit situated between the myocardium and visceral pericardium [4]. Unlike other fat stores, it is metabolically active and contributes to myocardial lipid metabolism, inflammatory mediator secretion, and coronary artery function [5]. The distinct characteristics of EAT stem from its specialized transcriptome, which dynamically responds to cardiovascular conditions such as coronary artery disease (CAD), atrial fibrillation (AF), and heart failure (HF) [6, 7, 8]. Advances in molecular biology have enabled detailed profiling of EAT gene expression, revealing roles in inflammation, lipid metabolism, and energy regulation [9, 10]. Notably, the transcriptomic landscape of EAT is altered by systemic and local inflammation induced by CPB, suggesting a complex interplay between EAT and postoperative inflammatory pathways [11]. Emerging evidence further indicates that EAT can serve as a local source of inflammatory mediators that act through paracrine and endocrine mechanisms [12].
The objective of this study is to explore and uncover the biomolecular changes induced by CPB in EAT and evaluate their potential contribution to perioperative inflammation and myocardial injury, with the goal of identifying therapeutic targets to mitigate these adverse effects. From each article, this paper will seek to identify the top biomarkers implicated in post-CPB systemic and cardiac inflammation. Understanding these changes may provide insights on the pathophysiological mechanisms underlying CPB-induced inflammation and its associated cardiac effects. Furthermore, targeting these pathways may provide a therapeutic strategy to mitigate the adverse effects associated with the inflammatory response in these patients [13].
This review was conducted in compliance with the 2020 Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) statement [14]. Electronic searches were conducted in MEDLINE and EMBASE with keywords: [Epicardial] AND [(cardiopulmonary bypass)] AND [(cardiac surgery OR heart surgery)] from database inception to January 2025. All primary research papers published in English that reported molecular changes within EAT before and after cardiac surgery with CPB were included. Pediatric patients, heart-transplant cases, and minimally invasive cardiac procedures that did not involve CPB were excluded. Abstracts, editorials, case reports, and reviews were also excluded. The following was extracted from each study: study design, study size, patient baseline characteristics, location of EAT extraction, EAT preparation mode, time points of EAT collection, location of subcutaneous adipose tissue (SAT) extraction, SAT preparation mode, time points of SAT collection, molecule/gene studied, mode of molecule/gene analysis, and molecular change observed following surgery. Primary outcomes included changes in local protein levels and/or mRNA expression in EAT following surgery. Secondary outcomes included the subsequent measurements of protein level changes or mRNA expression changes within SAT and serum. Studies which did not measure biomarker levels in EAT both before and after CPB were not included since there was no baseline to compare the effects of CPB against. References were uploaded to Covidence Systematic Reviews software and duplicates were removed. Two screeners (J.S) and (H.Z) independently screened studies to determine their eligibility.
The search identified 1450 studies for screening, of which 85 were eligible for full-text review. After assessment, 80 studies were excluded for the following reasons: absence of pre- and postoperative biomarker data (n = 53), publication as journal or congress abstracts (n = 18), lack of EAT collection (n = 5), non-cardiac surgical procedures (n = 2), and non-English language (n = 2). Ultimately, five studies were included in this review (Fig. 1) [15, 16, 17, 18, 19]. All analyzed and stored paired EAT and SAT samples before and after CPB and adhered to the Declaration of Helsinki. Study characteristics are summarized in Table 1 (Ref. [15, 16, 17, 18, 19]), and key findings in Table 2 (Ref. [15, 16, 17, 18, 19]).
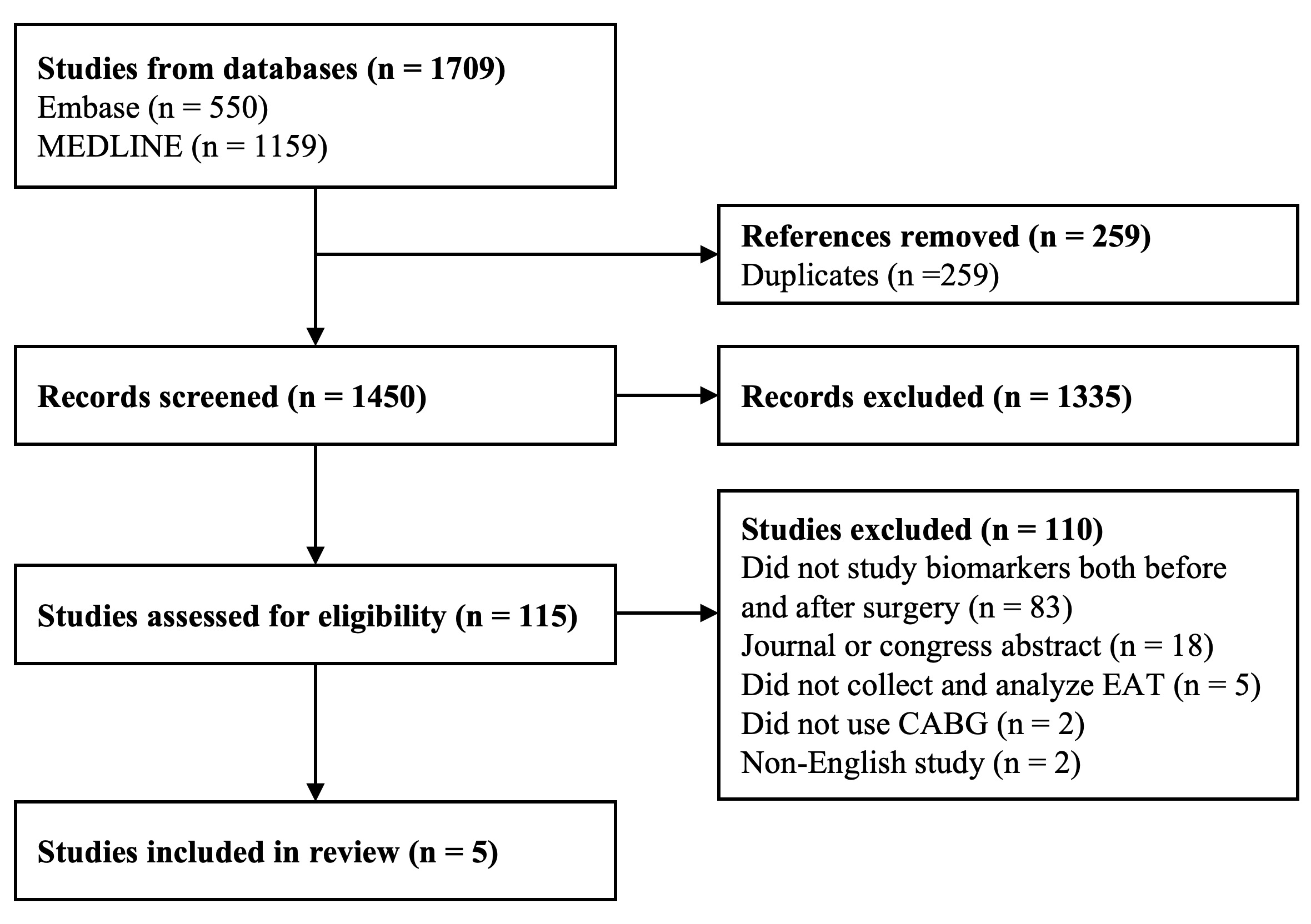 Fig. 1.
Fig. 1.
PRISMA flow diagram showing the selection process for included studies. PRISMA, Preferred Reporting Items for Systematic Reviews and Meta-Analyses.
| Study | Investigation | Stratification | Patient demographics | Surgery type | Sample place | Site of EAT extraction | Measurement of EAT expression | Biomarkers |
| Kotulák et al., 2011 [15] | Pre- and Postoperative Fibroblast Growth Factor- 21 Level | No stratification | n = 15 age: 66 |
CABG | EAT, SAT, and serum | Not indicated | mRNA level measurement: qRT-PCR | TNF-alpha, IL-6, IL-8, MCP-1, C-reactive protein, glucose, insulin, FGF-21 |
| male: 100% | ||||||||
| BMI |
Protein level measurement: Western blotting | |||||||
| Mach et al., 2017 [16] | Pre- and Postoperative cytokines level | No stratification | n = 77 age: 67.68 |
CABG | EAT and SAT | Near the proximal left and right coronary arteries | Protein level measurement by ELISA and bicinchoninic acid assay | TNF- |
| female: 36%, male: 64% | ||||||||
| BMI: 29.02 kg/m2 | ||||||||
| Matloch et al., 2022 [17] | Pre- and Postoperative Epicardial CTRP3 mRNA Levels in Patients with Type 2 Diabetes | -CAD-T2DM | n = 11 Age: 59.7 |
CABG | EAT, SAT, and serum | Anterior interventricular sulcus or the right margin of the heart | mRNA level measurement by qRT-PCR | C1q TNF-related protein 3 (CTRP3), TNF- |
| BMI: 27.1 | ||||||||
| +CAD-T2DM | n = 14 Age: 69.3 | |||||||
| BMI: 28.3 | ||||||||
| +CAD+T2DM | n = 9 Age: 67.0 | |||||||
| BMI: 30.4 | ||||||||
| Kratochvílová et al., 2021 [18] | Pre- and Postoperative Mitochondrial and Endoplasmic Reticulum Stress Genes Level | CAD | n = 27 age: 67.6 |
CABG and/or valve replacement | EAT and SAT | Anterior interventricular sulcus or the right margin of the heart | mRNA level measurement by qRT-PCR | TNF- |
| female: 19%, male: 81% | ||||||||
| BMI: 28.6 | ||||||||
| No CAD | n = 11 age: 59.8 | |||||||
| female: 27%, male: 73% | ||||||||
| BMI: 27.1 | ||||||||
| Kotulak et al., 2014 [19] | Pre- and Postoperative A-FABP Level | +T2DM | n = 11 age: 62.0 |
CABG, aortic valve replacement (AVR), and mitral valve replacement (MVR) | EAT, SAT, and serum | Not indicated | mRNA level measurement by qRT-PCR | A-FABP, insulin, glucose, IL-6, IL-8, MCP-1, and hsCRP |
| female: 0%, male: 100% | ||||||||
| BMI: 31.1 | ||||||||
| -T2DM | n = 20 age: 64.7 |
Protein level measurements by ELISA, high sensitive assay, radioimmunoassay, and LINCOplex | ||||||
| female: 0%, male: 100% | ||||||||
| BMI: 29.4 |
CABG, coronary artery bypass graft; AVR, aortic valve replacement; MVR, mitral valve replacement; EAT, epicardial adipose tissue; SAT, subcutaneous adipose tissue; ER, endoplasmic reticulum; BMI, body mass index; qRT-PCR, quantitative reverse transcription polymerase chain reaction; A-FABP, adipocyte fatty acid-binding protein; CTRP3, C1q/TNF-related protein 3; hsCRP, high-sensitivity C-reactive protein.
| Study | Investigation | Molecule | Mode of measurement in EAT and SAT | Stratification | EAT | SAT | Serum |
| Kotulák et al., 2011 [15] | Pre- and postoperative Fibroblast Growth Factor-21 level | FGF21 | mRNA and Protein | Pooled | ↑* | = | ↑* until 24 h post-surgery |
| Mach et al., 2017 [16] | Pre- and postoperative cytokines level | leptin | Protein | Pooled | ↓* | = | N.S. |
| adiponectin | Protein | Pooled | ↓* | = | |||
| TNF- |
Protein | Pooled | ↓* | = | |||
| A-FABP | Protein | Pooled | ↓* | = | |||
| IL-6 | Protein | Pooled | ↓ | ↓* | |||
| Matloch et al., 2022 [17] | Pre- and postoperative epicardial CTRP3 and inflammatory cytokine levels in patients with Type 2 Diabetes | IL-6 | mRNA | -CAD-T2DM | ↑* | ↑* | ↑ |
| +CAD-T2DM | ↑* | ↑* | ↑ | ||||
| +CAD+T2DM | ↑* | ↑* | ↑* | ||||
| TNF- |
mRNA | -CAD-T2DM | ↑* | ↑* | ↑* | ||
| +CAD-T2DM | ↑ | ↑* | ↑ | ||||
| +CAD+T2DM | ↑ | ↑* | ↑* | ||||
| CTRP3 | mRNA | -CAD-T2DM | ↑ | = | ↓* | ||
| +CAD-T2DM | ↑ | = | ↓* | ||||
| +CAD+T2DM | ↑ | = | ↓* | ||||
| adiponectin | N.A. | -CAD-T2DM | N.S. | ↓ | |||
| +CAD-T2DM | ↓ | ||||||
| +CAD+T2DM | ↓* | ||||||
| leptin | N.A. | -CAD-T2DM | ↓* | ||||
| +CAD-T2DM | ↑ | ||||||
| +CAD+T2DM | ↑ | ||||||
| Kratochvílová et al., 2021 [18] | Pre- and postoperative mitochondrial and endoplasmic reticulum stress genes level | MT-ND5 | mRNA | no CAD | ↓* | ↓ | N.S. |
| CAD | ↓* | ↓* | |||||
| COX4/1 | mRNA | no CAD | ↓ | ↓ | |||
| CAD | ↑ | ↑ | |||||
| CS | mRNA | no CAD | ↓* | ↓ | |||
| CAD | ↓* | ↑ | |||||
| NDUFA12 | mRNA | no CAD | ↓* | ↓ | |||
| CAD | ↓* | ↑ | |||||
| CYC1 | mRNA | no CAD | ↓ | ↓ | |||
| CAD | ↑ | ↑ | |||||
| SDHA | mRNA | no CAD | ↓ | ↓ | |||
| CAD | ↑ | ↑ | |||||
| HSPA5 | mRNA | no CAD | ↑ | ↑ | |||
| CAD | ↑* | ↑ | |||||
| DDIT3 | mRNA | no CAD | ↑ | ↓ | |||
| CAD | ↑ | = | |||||
| ATF4 | mRNA | no CAD | ↑* | ↑ | |||
| CAD | ↑* | ↑ | |||||
| ATF6 | mRNA | no CAD | ↑ | ↑ | |||
| CAD | ↑* | ↑ | |||||
| Kotulak et al., 2014 [19] | Pre- and postoperative A-FABP level | A-FABP | mRNA | +T2DM | ↓ | = | ↑* until 6 h post-surgery |
| -T2DM | ↑ | ↓ | ↑* until 24 h post-surgery | ||||
| IL-6 | N.A. | +T2DM | N.S. | ↑* until 24 h post-surgery | |||
| -T2DM | ↑* until 48 h post-surgery | ||||||
“*”: indicates statistical significance (p
Kotulák et al. (2011) [15] measured both serum protein and EAT/SAT mRNA levels. All other studies included both serum and measured mRNA levels of their protein of interest in EAT and SAT [16, 17, 18, 19]. All studies reported mRNA levels in normalized, relative units of gene expression. Since this study aimed to compare changes in expression levels, the quantification and analytical methods used by the selected studies were both appropriate and sufficient.
Methodologies for data acquisition in all five studies were similar, providing variable control and enabling the direct comparisons performed in this study. Patients fasted overnight before having surgery with CPB started between 7–8AM [15, 16, 17, 18, 19]. In all cases, blood samples were drawn before administration of anesthesia and at the end of surgery, with serum isolated by centrifugation. EAT was harvested from the anterior interventricular sulcus or right margin of the heart, and SAT from the sternotomy site in the thoracic region [15, 16, 17, 18, 19]. Both tissue samples were collected at the start and end of surgery in all cases.
Several biomarkers were identified with suspected roles in post-CPB inflammation and adverse cardiac remodeling. Fibroblast growth factor 21 (FGF-21) was included as a potential response marker to CPB-induced cardiac stress [15]. Interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-alpha) are classic pro-inflammatory cytokines known to have a broad range of activity, including in post-CPB cardiac remodeling and heart failure [16, 17, 19]. The adipokines leptin and adipocyte fatty acid binding protein (A-FABP), and adiponectin and C1q/TNF-related protein 3 (CTRP3), are also known to have pro- and anti-inflammatory effects, respectively, in cardiovascular diseases (CVDs) [17, 19]. Lastly, a panel of mitochondrial and ER stress genes were also included as markers of interest due to their role in contributing to cardiac dysfunction and heart failure [18].
FGF-21 mRNA expression increased significantly (p = 0.016) within EAT after CPB surgery but not within SAT [15]. Serum FGF-21 concentrations were found to increase over baseline 6 hours post-surgery, which reverted to baseline levels 96 hours post-surgery [15].
IL-6 protein levels in both EAT (N.S) and SAT (p = 0.0209) decreased
post-surgery [16], and its mRNA expression increased (p
TNF-alpha protein levels increased significantly in EAT (p
CTRP3 mRNA expression increased in EAT, albeit nonsignificant, in all patients
(p
EAT protein levels of leptin, adiponectin, and A-FABP decreased significantly
(p = 0.001, p
There was generally a post-surgical decrease in mRNA expression of all
mitochondrial ETC genes in EAT in patients with or without CAD (p
There was generally a post-surgical increase in mRNA expression of ER stress
genes in both EAT and SAT (p
This systematic review highlights the molecular changes in the EAT of patients who underwent cardiac surgery with CPB. Our findings indicate a marked increase in inflammatory markers, notably TNF-alpha and IL-6, in EAT and in systemic circulation post-CPB. This amplified inflammatory response aligns with previous studies that have implicated TNF-alpha and IL-6 in adverse cardiac remodeling and heart failure [20]. Elevated TNF-alpha and IL-6 levels have also been shown to induce cardiomyocyte apoptosis, myocardial fibrosis, and impaired cardiac contractility, contributing to the development of HF [21]. These cytokines also activate systemic inflammatory pathways, exacerbating post-surgical cardiac dysfunction and remodeling [22].
Patients undergoing cardiac surgery often present with comorbidities such as CAD and T2DM, both of which are characterized by chronic low-grade inflammation. This pre-existing pro-inflammatory milieu may influence baseline biomarker expression within EAT and amplify the perioperative cytokine response to CPB [1, 2]. Consequently, observed elevations in IL-6 and TNF-alpha post-CPB likely reflect an interplay between chronic metabolic inflammation and acute surgical stress. This interaction may partly explain the inter-patient variability in inflammatory biomarker trajectories and underscores the importance of accounting for baseline inflammatory status in future studies.
The studies included also showed that mRNA expression of IL-6 increased in both EAT and SAT [17]. Despite this increase, local protein levels of IL-6 decreased, while serum levels rose [17, 19]. mRNA expression of TNF-alpha also increased in both EAT and SAT [17]. However, local protein levels decreased only in EAT [16], with an accompanying rise in serum levels [17]. These findings suggest that cardiac adipose tissue is biologically active. Evidently, EAT secretes proteins that either remain local and exert their direct effect on the cardiac muscle or reach circulation for a systemic effect through pericardial fluid, lymphatic drainage, or venous return [7, 23, 24].
Our review also showed a post-surgical decline in local EAT levels of adipokines (leptin, adiponectin, and A-FABP) with no changes in SAT. Serum leptin decreased in healthy patients but increased in those with comorbidities [16]. Moreover, adiponectin decreased [17], and A-FABP increased in circulation across all patients [19]. In healthy individuals, leptin regulates appetite and energy balance, but also contributes to cardiovascular health. Elevated leptin levels are linked to obesity and CVDs due to its pro-inflammatory effects [25]. Adiponectin, known for its anti-inflammatory and insulin-sensitizing properties, conversely protects against atherosclerosis and improves endothelial function [25]. Lastly, A-FABP - is crucial in lipid metabolism and glucose homeostasis, with its high levels associated with an increased risk of metabolic syndrome and CVDs due to its role in promoting inflammation and insulin resistance [26].
The observed post-surgical changes in these adipokines may significantly impact metabolic and cardiovascular outcomes in patients undergoing CPB. The variable response of adipokines in the context of existing cardiac diseases underscores a multifaceted relationship between adipokine regulation and cardiovascular health. For example, changes in leptin signaling are associated with cardiac dysfunction [27]. Conversely, adiponectin is cardioprotective, and its reduction post-CPB suggests a connection to induced cardiac stress [28]. A-FABP contributes to insulin resistance and metabolic regulation such as lipid trafficking. Increased A-FABP levels are linked to a higher prevalence of adverse cardiac events, diastolic dysfunction, and left ventricular hypertrophy in diabetic patients without previous CVD [29, 30].
Beyond their pro-inflammatory effects, recent literature also emphasizes the dual nature of EAT as both a potential mediator of inflammation and a source of cardioprotective factors [24, 26, 31]. Under physiological conditions, EAT secretes anti-inflammatory and vaso-protective adipokines; such as adiponectin and adrenomedullin, that help maintain endothelial function and myocardial metabolism [5, 6, 28]. However, surgical stress, ischemia, or metabolic dysfunction may shift this balance toward a pro-inflammatory phenotype, reducing its protective capacity. Understanding this bidirectional role of EAT is crucial for interpreting perioperative molecular findings and identifying therapeutic strategies that preserve its protective signaling after cardiopulmonary bypass.
Furthermore, there was a consistent reduction in mRNA expression levels of all mitochondrial ETC genes in EAT, but not in SAT [18]. ER stress genes also appeared to non-specifically decrease in both EAT and SAT; however, some markers (ATF4, DDIT3, ATF6, HSPA5) were differentially upregulated in patients with comorbidities such as CAD [18]. Under normal conditions, mitochondrial ETC genes are crucial for energy production through oxidative phosphorylation [32]. Their dysfunction can lead to metabolic disturbances and CVDs due to impaired ATP synthesis and increased production of reactive oxygen species (ROS) [32]. This oxidative stress can compromise cellular structures and exacerbate metabolic stress, particularly in patients with CAD [32]. Therefore, the downregulation of ETC genes in EAT post-CPB points to impaired mitochondrial function, potentially underpinning the observed metabolic disturbances. The ER stress genes, including ATF4, DDIT3, ATF6, and HSPA5, help maintain protein homeostasis and manage cell stress. Dysregulation of these genes activates the unfolded protein response (UPR), which aims to restore ER function by enhancing protein folding capacity and degrading misfolded proteins [33]. However, prolonged or severe ER stress can lead to apoptosis, inflammation, and endothelial dysfunction, contributing to atherosclerosis and heart failure [32, 34]. The increase of these ER stress genes in both EAT and SAT underscores the cellular stress response to CPB, with a more pronounced effect in EAT.
In addition to its metabolic and inflammatory roles, EAT also harbors stromal and stem cell populations with paracrine and regenerative potential [24, 29]. These cells contribute to myocardial repair through the secretion of growth factors, cytokines, and extracellular vesicles that support angiogenesis and cardiomyocyte survival. Emerging evidence suggests that biomolecules such as FGF-21 and adiponectin; both altered following CPB, may modulate the activity of these stromal cells, influencing their reparative and immunomodulatory functions [15, 28, 29, 30]. The observed molecular changes in this review may therefore not only reflect inflammation and metabolic stress but also adaptive responses within the EAT stromal cell niche that influence postoperative tissue remodeling.
These molecular shifts hint at potential pathways through which CPB can influence cellular and myocardial health. Maintaining mitochondrial and ER integrity is crucial, as these systems play key roles in cellular energy metabolism and stress response. Strategies that enhance mitochondrial biogenesis, reduce oxidative stress, and support ER function could potentially improve postoperative outcomes in CPB patients [35, 36]. The proposed pathophysiology of the biomarkers discussed in this section are presented in Fig. 2.
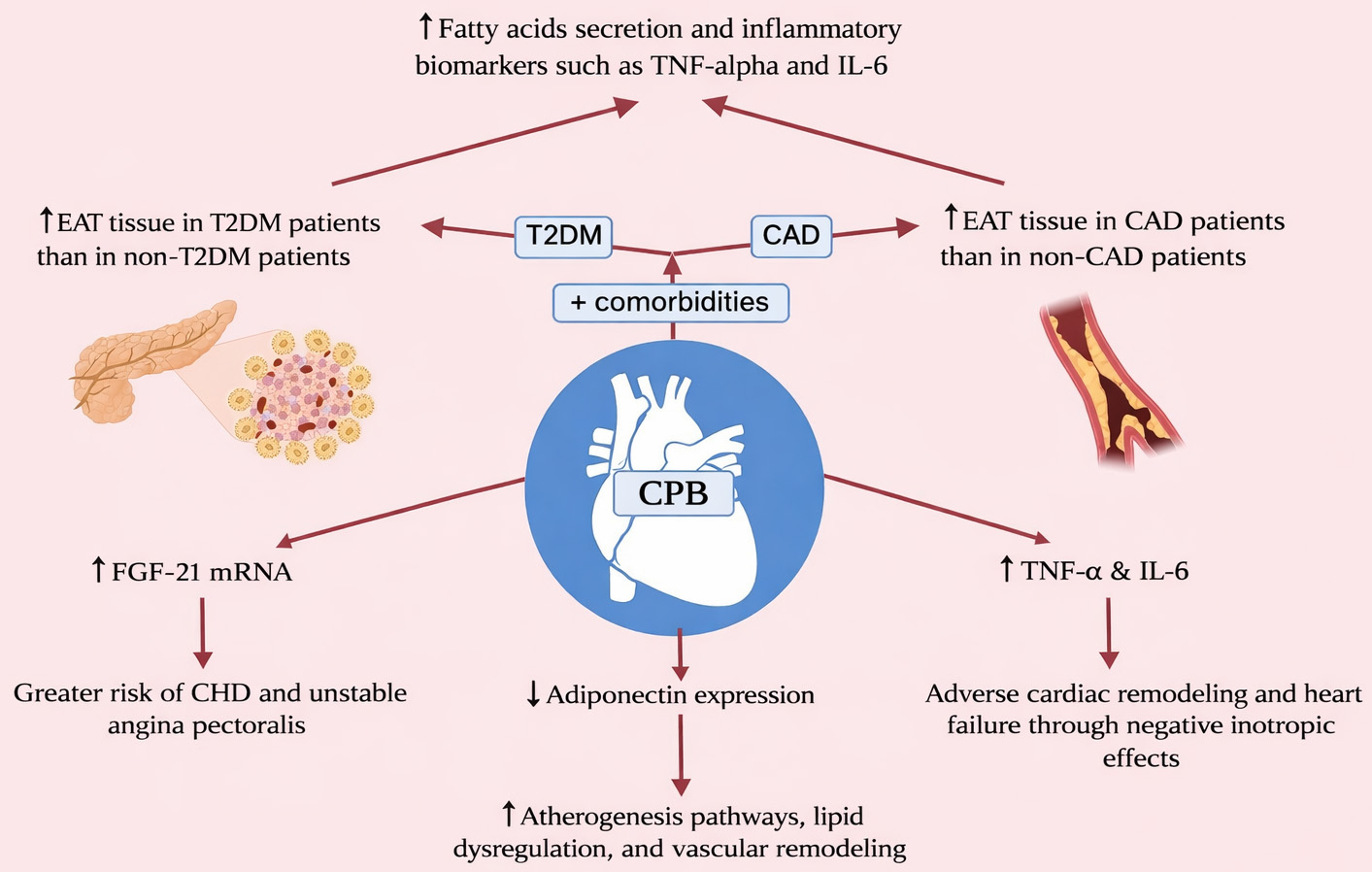 Fig. 2.
Fig. 2.
Proposed pathophysiology of inflammatory markers.
The distinct response to CPB in cardiac adipose tissue, particularly concerning inflammatory markers, adipokines, and stress-related genes, inform the development of potential therapeutic targets for enhancing postoperative recovery [37]. Pharmacological interventions, such as anti-inflammatory agents, may mitigate the observed post-CPB inflammatory spike of TNF-alpha and IL-6, resulting in potential attenuation of the systemic inflammatory response associated with CPB [38, 39]. Additionally, leveraging thiazolidinediones (TZDs) to elevate adiponectin levels could safeguard against post-surgical cardiac complications due to their cardioprotective properties [30]. Strategies promoting mitochondrial health and alleviating ER stress also suggest promise. For example, compounds enhancing mitochondrial biogenesis or functionality, such as resveratrol or coenzyme Q10 have been shown to counteract age-related mitochondrial dysfunction by counterbalancing differential mitochondrial gene expression [40]. These compounds may similarly mitigate the surgery-induced decline in EAT’s mitochondrial gene expression. Furthermore, reperfusion injury may be significant to consider, as mRNA expression of FGF-21 in EAT significantly increases after CPB [15]. Since FGF-21 plays a role in protecting myocardial tissue against ischemic damage, it may be beneficial to prophylactically administer anti-oxidative pharmaceutical agents [41]. An animal study conducted by Salameh et al. (2020) [42] showed that the anti-oxidants epigallocatechin-3-gallate and minocycline significantly lowered myocardial damage post-CPB. In addition to pharmaceutical prophylaxis, there are animal studies demonstrating that terminal leukocyte-depleted lung reperfusion may reduce ischemic injury post CPB [43].
This review is limited by the small number of eligible studies (n = 5) and the observational design of all included investigations. The heterogeneity of study populations, biomarker targets, and analytical methodologies precluded the performance of a quantitative meta-analysis. Instead, we synthesized findings narratively in accordance with PRISMA-ScR guidelines, emphasizing consistency in molecular trends rather than statistical aggregation. Furthermore, the absence of standardized biomarker measurement techniques across studies may have introduced variability in reported effect magnitudes and limits the generalizability of results. Future prospective and mechanistic studies with harmonized protocols are needed to validate and expand upon these findings.
Our findings suggest that CPB can induce significant biomolecular changes in EAT which may potentially contribute to perioperative inflammatory changes and myocardial injury. Targeting these changes may provide a potential therapeutic strategy to mitigate these adverse effects in this group of patients.
AF, Atrial Fibrillation; A-FABP, Adipocyte Fatty Acid-Binding Protein; ATF4, Activating Transcription Factor 4; ATF6, Activating Transcription Factor 6; CAD, Coronary Artery Disease; CPB, Cardiopulmonary Bypass; CTRP3, C1q/TNF-Related Protein 3; DDIT3, DNA Damage Inducible Transcript 3; EAT, Epicardial Adipose Tissue; ER, Endoplasmic Reticulum; ETC, Electron Transport Chain; FABP4, Fatty Acid-Binding Protein 4; FGF-21, Fibroblast Growth Factor 21; HF, Heart Failure; HSPA5, Heat Shock Protein A5; IL-6, Interleukin 6; mRNA, Messenger Ribonucleic Acid; PRISMA, Preferred Reporting Items for Systematic Reviews and Meta-Analyses; ROS, Reactive Oxygen Species; SAT, Subcutaneous Adipose Tissue; TNF-alpha, Tumor Necrosis Factor Alpha; TZDs, Thiazolidinediones; UPR, Unfolded Protein Response.
All data is available upon request to the corresponding author.
AS, YD, HLZ, AM, JS, YAO, MED contributed to the conception and editing of the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All included studies conform to the standards laid in the 1964 Declaration of Helsinki.
Not applicable.
This research received no external funding.
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/HSF50005.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.