











 , Bindu Jayashankar 2,*
, Bindu Jayashankar 2,* , Raghu Nataraj 1,*
, Raghu Nataraj 1,*1 Division of Molecular Biology, School of Life Sciences, Jagadguru Shri Shivaratreeshwara (JSS) Academy of Higher Education and Research, 570015 Mysore, Karnataka, India
2 Department of Biotechnology, SJCE, JSS Science & Technology University, 570006 Mysore, Karnataka, India
Abstract
The hypothalamic-pituitary-ovarian (HPO) axis is complexly dysregulated in female infertility, and this dysfunction is further exacerbated by metabolic and nutritional abnormalities. Diet and nutrient-derived bioactives significantly influence neuroendocrine factors such as insulin signalling, gonadotropin-releasing hormone (GnRH) pulsatility, and inflammatory tone. Insight into the intersection of these molecular and nutritional pathways may reveal new opportunities for restoring fertility. This review aims to integrate the neuroendocrine, metabolic, and nutritional mechanisms governing female fertility while identifying convergent molecular hubs as potential therapeutic targets.
A structured search of English-language studies published between 2010 and 2025 was conducted using PubMed, Scopus, Web of Science, and Google Scholar. After screening 186 research and review articles, 92 that met the inclusion criteria were included. Data on neuroendocrine regulation, signalling pathways, and nutritional interventions were extracted. Additionally, 24 nutrient-responsive and reproductive regulatory genes with a minimum interaction confidence score of 0.7 were included in a targeted in-silico protein-protein interaction (PPI) network analysis using STRING v12.0 (Homo sapiens). Kyoto Encyclopedia of Genes and Genomes (KEGG) was utilized to evaluate pathway enrichment.
The integrated synthesis shows that phosphoinositide 3-kinase/protein kinase B/mammalian target of rapamycin (PI3K/AKT/mTOR), nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), and sirtuin 1 (SIRT1) signalling directly affect GnRH neuronal activity, ovarian steroidogenesis, and oocyte competence. In-silico analysis identified major central hubs connecting metabolic and neuroendocrine regulation, including AKT1, mTOR, estrogen receptor 1 (ESR1), SIRT1, nuclear factor kappa B subunit 1 (NFKB1), and peroxisome proliferator-activated receptor gamma (PPARG). Nutraceuticals that consistently targeted these hubs, including myo-inositol, vitamin D, omega-3 polyunsaturated fatty acids (PUFAs), resveratrol, curcumin, and quercetin, improved insulin sensitivity, decreased oxidative stress, and restored normal gonadotropin ratios. The evidence supports a multi-target, nutrition-based approach to fertility management.
The neuroendocrine and molecular regulators of female fertility are measurable targets for nutritional and nutraceutical interventions. Focusing on common signalling hubs identified in the literature and in-silico mapping can facilitate the development of standardized, artificial intelligence (AI)-guided nutraceutical formulations. Future research should incorporate biomarker-based endpoints to translate these mechanistic insights into therapeutic benefit.
Keywords
- female infertility
- nutrition
- neuroendocrine regulation
- hypothalamic-pituitary-ovarian axis
- PI3K/AKT/mTOR pathway
- nutraceuticals
- in silico analysis
- PCOS
- molecular signaling
- reproductive health
Female infertility, defined as the inability to achieve pregnancy after 12 months or more of unprotected intercourse, affects a substantial proportion of reproductive-aged women worldwide and has a significant psychosocial and economic impact on individuals and society. Infertility is estimated to affect 10–15% of couples worldwide, with female-related factors accounting for nearly half of these cases. The World Health Organization defines infertility as a disease of the reproductive system and a public health issue, especially in low- and middle-income countries, where diagnostic and treatment options are limited. In addition to biological causes, psychosocial stressors and poor nutrition frequently exacerbate infertility, further highlighting its complex and biopsychological nature. Conditions like ovulatory dysfunction, tubal disease and endocrine abnormalities are major contributors to female infertility [1, 2, 3].
At the molecular level, the female reproductive system is controlled by tightly regulated neuroendocrine signals, particularly the hypothalamic-pituitary-ovarian (HPO) axis, which ensures timing of gonadotropin-releasing hormone (GnRH), luteinizing hormone (LH)/follicle-stimulating hormone (FSH) release, as well as ovarian steroidogenesis. Disruption of this axis, whether due to metabolic, hormonal, or neurogenic factors, underlies many forms of female infertility (Fig. 1) [4].
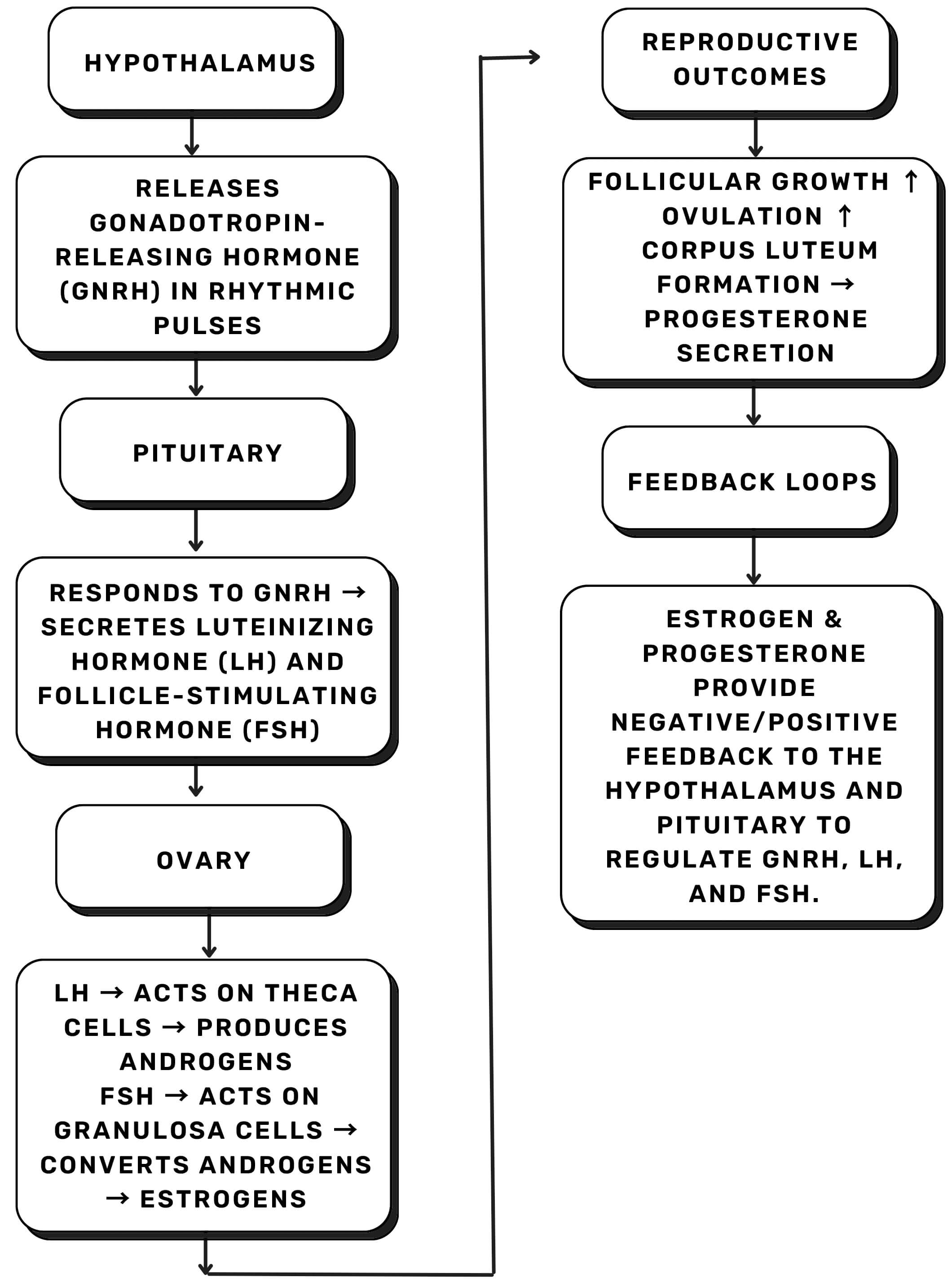 Fig. 1.
Fig. 1.
Simplified schematic of the hypothalamic-pituitary-ovarian (HPO) axis. Pulsatile GnRH release from the hypothalamus stimulates pituitary release of LH and FSH, which act on ovarian theca and granulosa cells to regulate steroidogenesis, ovulation, and luteal function. Estrogen and progesterone provide feedback to maintain hormonal balance. Abbreviations: GnRH, gonadotropin-releasing hormone; LH, luteinizing hormone; FSH, follicle-stimulating hormone. The Figs. 1–8 were created by the author using Canva (Cloud-based version, Canva Pty Ltd, Sydney, NSW, Australia).
In recent years, it has become clear that nutritional status and dietary patterns have a profound impact on female fertility through metabolic, endocrine, and neuroendocrine mechanisms. For example, diets high in refined carbohydrates, trans fats, and low in micronutrients have been shown to reduce fecundability [5].
Although research into neuroendocrine regulation and nutritional influences on fertility has advanced, a significant gap remains: an integrative understanding of how nutrition interacts with neuroendocrine signaling at the molecular level, especially in disorders such as polycystic ovary syndrome (PCOS) and other infertility etiologies [6].
This review aims to map the brain-to-ovary axis at the molecular level and to show how nutritional modulation can be applied in diagnostic and therapeutic strategies. In this article, we (i) review the neuroendocrine pathways of female reproduction, (ii) examine the molecular signaling cascades between the brain and ovary, (iii) assess how nutrition and nutraceuticals impact these pathways, (iv) discuss their diagnostic and therapeutic implications in female infertility, and (v) include a brief in silico network analysis (STRING) to illustrate the mechanistic link between nutritional modulation and female fertility.
Between October 2025 and November 2025, a comprehensive literature search was conducted using the databases PubMed, Scopus, Google Scholar, and Web of Science. The following keywords were searched: “female infertility”, “neuroendocrine regulation”, “hypothalamic-pituitary-ovarian axis”, “nutrition”, “nutraceuticals”, “molecular signaling”, “PCOS”, and “fertility therapy”. Terms were combined using the Boolean operators AND and OR. To ensure current relevance, only peer-reviewed English-language articles published between 2010 and 2025 were included.
The inclusion criteria comprised studies and reviews addressing: (a) neuroendocrine control of female reproduction; (b) molecular pathways connecting brain-ovary communication; (c) nutritional or nutraceutical interventions that affect reproductive or endocrine parameters; and (d) advancements in the diagnosis or treatment of female infertility. Articles that focused solely on male infertility, non-peer-reviewed sources, or animal models without translational relevance were excluded.
Key findings, nutrient mechanisms, molecular targets, and study type were among the data extracted. The data were grouped into thematic categories, including neuroendocrine mechanisms, molecular signaling pathways, nutritional modulation, and diagnostic or therapeutic perspectives. The findings were narratively synthesized to highlight integrative mechanisms and knowledge gaps synthesized.
To complement the literature-based synthesis, a targeted in silico analysis was performed to identify key molecular nodes linking neuroendocrine and nutritional pathways in female infertility.
The 24 genes included in the STRING analysis were selected based on a priori
criteria across five functional categories. First, genes with established
involvement in reproductive neuroendocrine regulation were included, specifically
kisspeptin receptor (KISS1R), gonadotropin-releasing hormone receptor (GNRHR),
estrogen receptor (ESR)1, and ESR2. Second, key regulators of ovarian steroidogenesis and follicular
function were incorporated, including cytochrome P450 aromatase (CYP19A1), steroidogenic acute regulatory protein (STAR), androgen receptor (AR), and 3beta-hydroxysteroid dehydrogenase2 (HSD3
Thus, the network represents the convergence point of four mechanistic domains: neuroendocrine regulation, steroidogenesis, metabolic-insulin signaling, and oxidative and inflammatory balance.
This framework reflects the scope of the present review and avoids the appearance of arbitrary selection.
The STRING protein-protein interaction (PPI) map revealed a tightly clustered core of highly connected nodes, including AKT1, mTOR, ESR1, PPARG, SIRT1, and NFKB1, which act as integration points for nutritional, metabolic, and hormonal signals (Fig. 2).
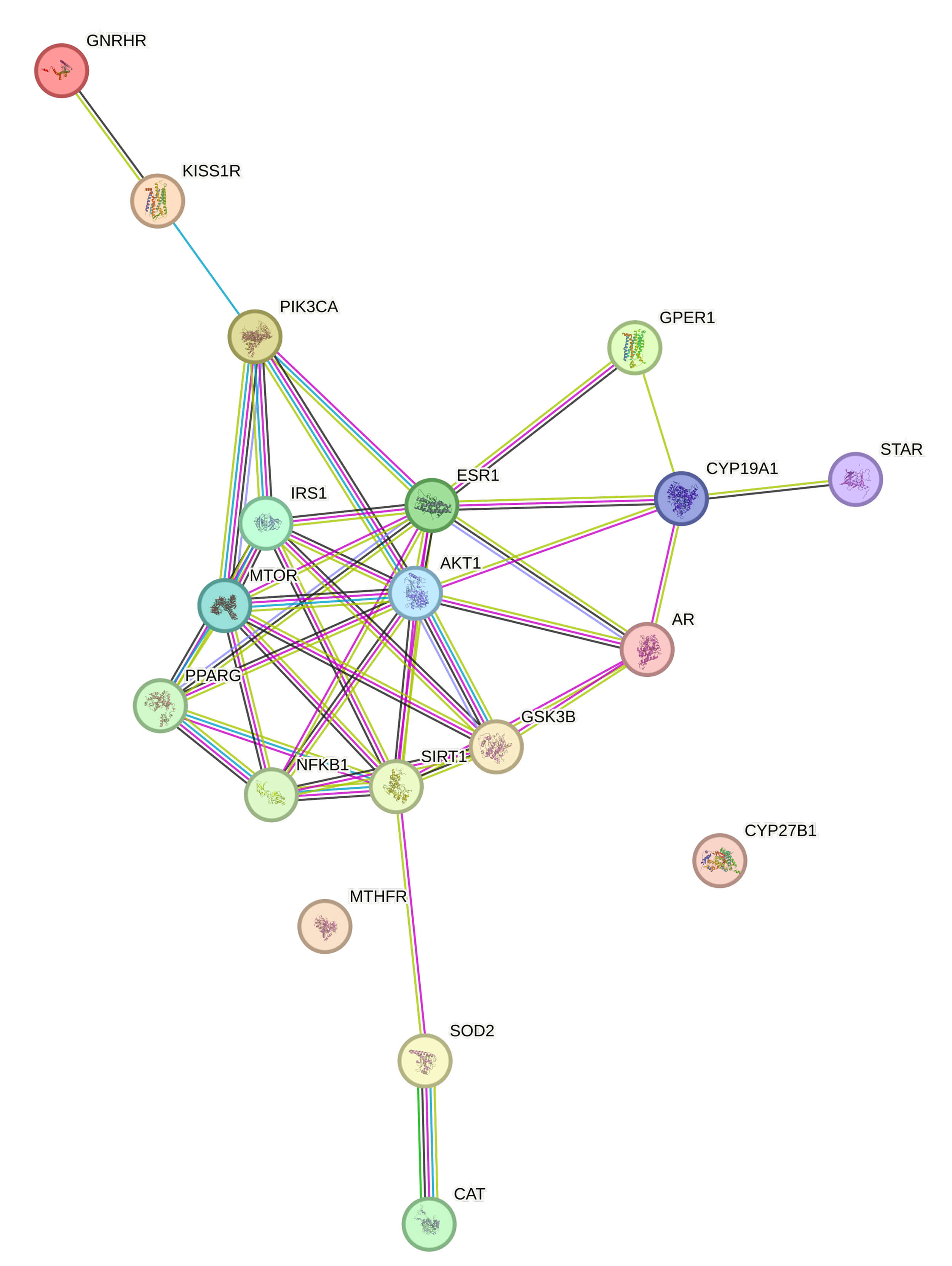 Fig. 2.
Fig. 2.
STRING-based protein-protein interaction (PPI) network depicting shared molecular targets of nutrient-responsive pathways involved in female fertility regulation. Central nodes (AKT1, mTOR, ESR1, PPARG, SIRT1, and NFKB1) represent key integration points linking metabolic, inflammatory, and neuroendocrine signaling. Abbreviations: AKT1, protein kinase B; mTOR, mammalian target of rapamycin; ESR1, estrogen receptor 1; PPARG, peroxisome proliferator-activated receptor gamma; SIRT1, sirtuin 1; NFKB1, nuclear factor kappa B subunit 1.
Physiological interpretation:
Peripheral nodes like CYP19A1, STAR, AR, and KISS1R/GNRHR provide anchoring between the PPI core and the physiological systems described earlier, namely steroidogenesis and hypothalamic-pituitary signaling.
Thus, the top pathways in Fig. 3 directly correspond to the reproductive tissues where these hubs are physiologically located:
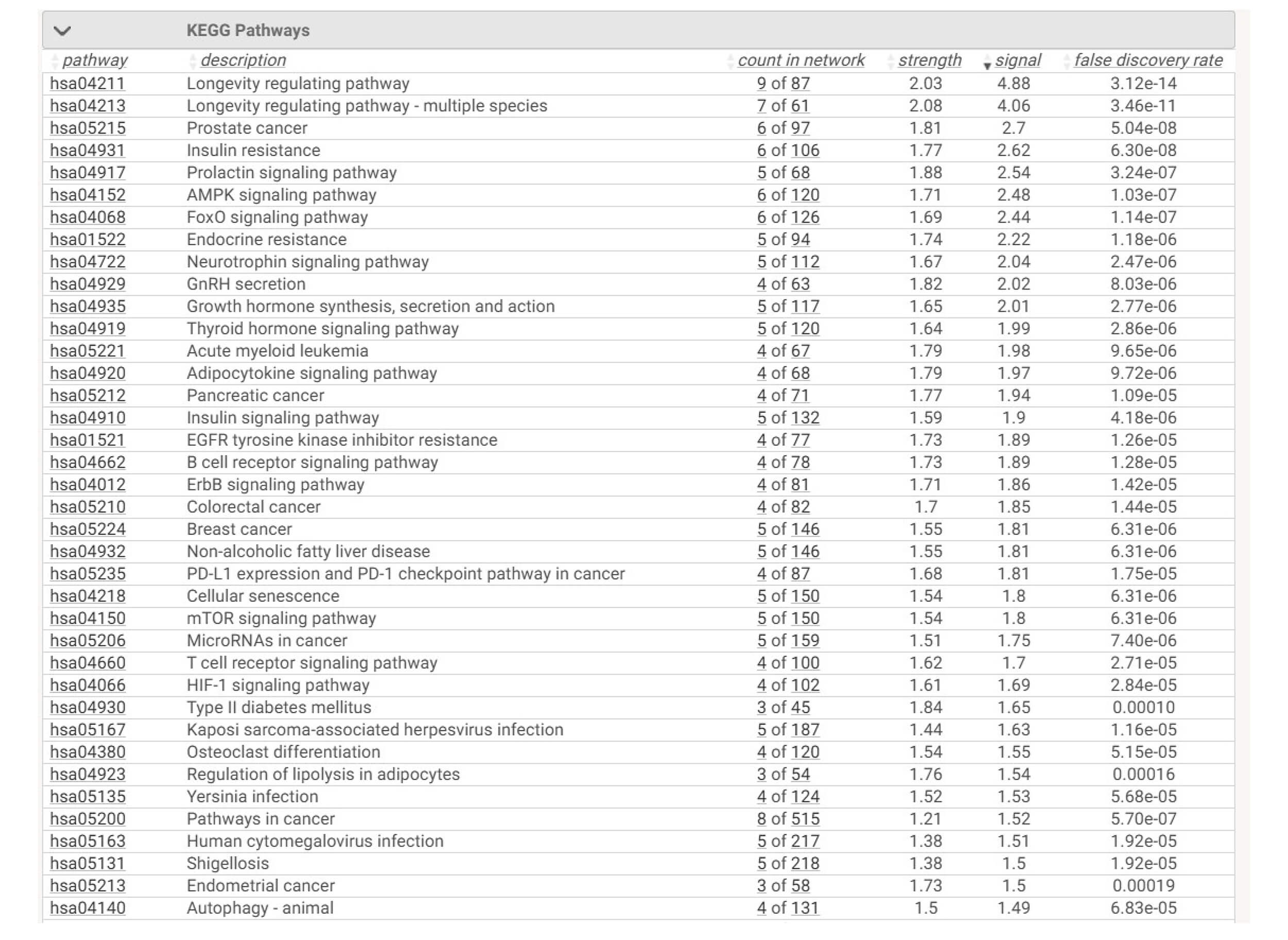 Fig. 3.
Fig. 3.
Enriched KEGG pathways derived from the nutrient-target PPI
network (confidence
The enriched pathways reflect the molecular architecture previously described:
3.1.3.1 Neuroendocrine Control
KISS1R–GNRHR interactions feed into PI3K/AKT and MAPK modules, showing how nutritional modulation of insulin and inflammatory status can alter GnRH pulsatility and gonadotropin output.
3.1.3.2 Ovarian Intracellular Signaling
mTOR, AKT1, ESR1, and NF-
3.1.3.3 Nutraceutical Mechanisms
Each major nutrient group maps directly onto a dominant hub:
Thus, the network reinforces that nutrient effects are not isolated but converge on shared mechanistic bottlenecks crucial for female fertility.
The combined experimental and in silico evidence suggests that:
This systems-level convergence helps explain why diverse interventions, including low-glycemic index (GI) diets, omega-3 enrichment, adequate vitamin D, antioxidant therapy, or inositol supplementation, consistently improve ovulatory function and IVF outcomes.
The following sections synthesize current evidence on neuroendocrine regulation, molecular signaling pathways, and nutritional modulation in female fertility.
The HPO axis is the central neuroendocrine pathway that drives female reproductive cycling by converting pulsatile hypothalamic GnRH into temporally regulated pituitary secretion of LH and FSH, which in turn control ovarian folliculogenesis, steroidogenesis, and ovulation [7].
Upstream neuropeptides that sense the hormonal and metabolic milieu regulate GnRH neuronal activity. Among these, kisspeptin neurons are the main gatekeepers of the reproductive hormones’ release. Kisspeptin signaling in the arcuate and preoptic hypothalamus potently stimulates GnRH release and encodes both negative and positive sex steroid feedback, which governs pulsatile vs surge modes of gonadotropin secretion. This kisspeptin-GnRH module is now recognized as a key neuroendocrine node that integrates metabolic, circadian, and steroid feedback to control reproductive function [8].
The anterior pituitary receives hypothalamic signals and converts neural signals into endocrine responses that directly affect the ovaries. Thus, pituitary gonadotropins act as the effector hormones of the HPO axis, where FSH promotes granulosa cell proliferation and the aromatase-dependent conversion of androgens to estradiol, whereas LH acts on theca cells and, later, on mural granulosa cells to promote androgen synthesis, follicular maturation, and the preovulatory surge that triggers oocyte release and luteinization. The interplay between follicular estradiol production and pituitary responsiveness controls follicle selection and the preovulatory surge [9].
Several nodes of the HPO axis are subject to tightly controlled feedback from ovarian steroids. Progesterone modulates the frequency of the GnRH pulse and primes the endometrium for implantation following ovulation, whereas estradiol exerts both positive and negative feedback on the amplitude of the GnRH pulse through arcuate circuitry and preoptic kisspeptin neurons. Reproductive readiness and regular cycle progression depend on these steroid feedback loops [7, 10].
In addition to gonadal feedback, stress-responsive systems have a significant impact on the reproductive axis. Stress hormones can alter reproductive signaling, as the hypothalamic-pituitary-adrenal (HPA) and HPO axes have reciprocal regulatory relationships. Psychological and physiological stressors primarily inhibit reproductive function through activation of the HPA axis and subsequent elevation of glucocorticoid levels. These factors reduce the release of LH and FSH secretion and interfere with folliculogenesis by impairing the activity of kisspeptin and GnRH neuronal activity and by increasing inhibitory signals like gonadotropin-inhibitory hormone (GnIH). Reduced gonadotropin secretion and altered ovarian steroidogenesis have been demonstrated in both acute and chronic stress paradigms [11].
HPO function is significantly altered by metabolic disturbances. Peripheral insulin resistance and hyperinsulinemia increase the ovarian theca cell androgen production and intensify luteinizing responses, which lead to anovulatory phenotypes such as PCOS. Concurrently, altered insulin signaling in the pituitary and hypothalamus disrupts GnRH and gonadotropin dynamics. Thus, insulin resistance links systemic metabolic state to ovarian steroidogenic output and follicular arrest [12].
Beyond insulin resistance, neuroendocrine fertility control is also influenced by dietary and adipose-derived cues. Obesity and adiposity disrupt neuroendocrine reproductive regulation by altering leptin signaling, increasing the peripheral aromatization of androgens to estrogens in adipose tissue, promoting chronic low-grade inflammation, and altering hypothalamic nutrient-sensing circuits. These changes impair follicular development, disrupt GnRH pulsatility, and reduce oocyte quality, all of which contribute to a decrease in fertility [13].
A series of intracellular molecular cascades converts external hormonal and
metabolic cues into precise cellular responses within reproductive tissues and
form the basis of this endocrine coordination. An intricately coordinated network
of intracellular signaling pathways connects neuroendocrine cues, metabolic
inputs, and ovarian responses to regulate female fertility (Fig. 4). These
pathways—PI3K/AKT/GSK3
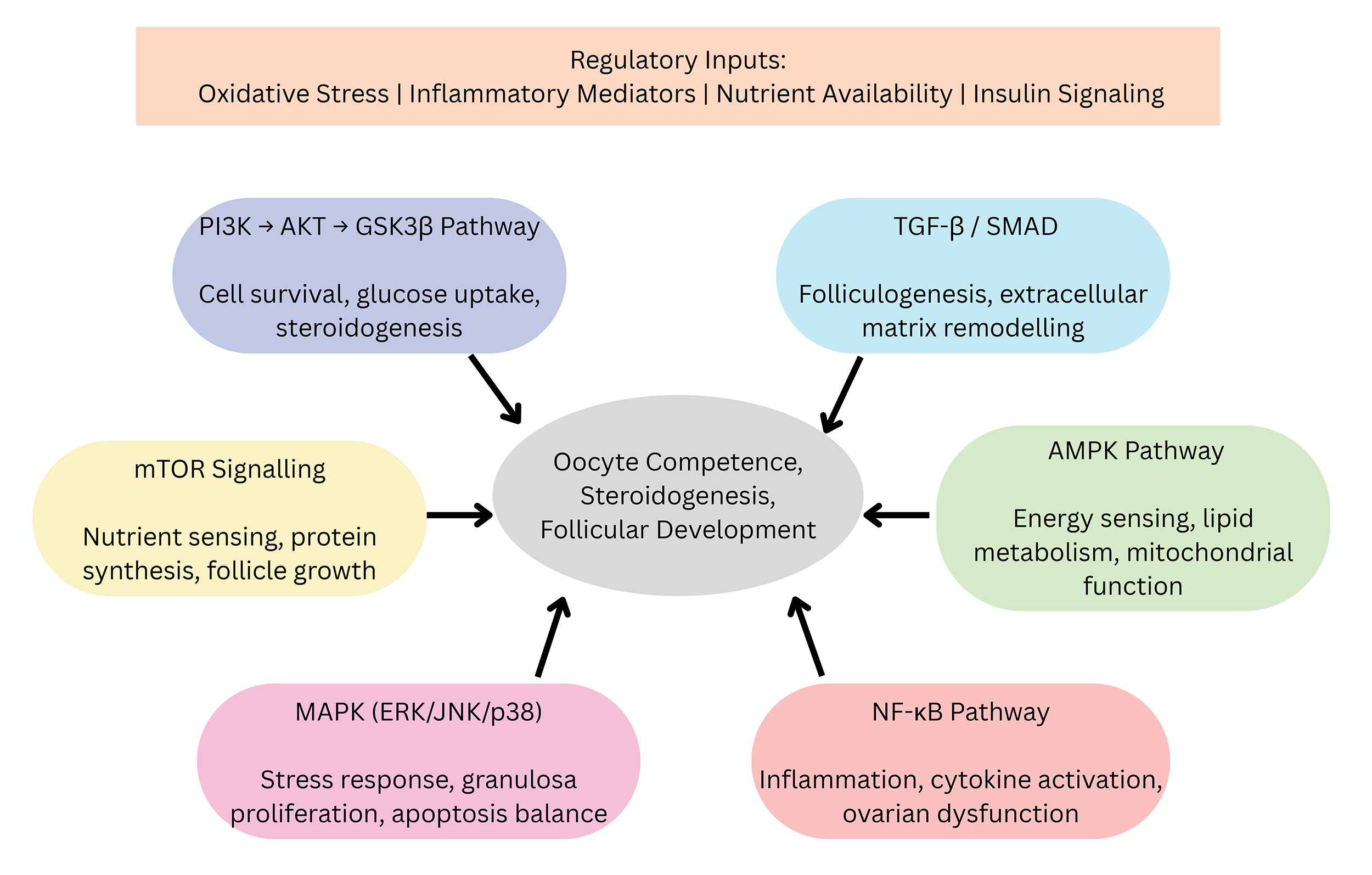 Fig. 4.
Fig. 4.
Integrated molecular pathways that control ovarian function.
Oxidative stress, inflammatory mediators, nutrient availability, and insulin
signaling act as upstream regulatory inputs that converge on six key
intracellular pathways—PI3K/AKT/GSK3
The effects of insulin, gonadotropins, and growth factors in reproductive
tissues are mediated by the crucial intracellular signaling cascade known as the
PI3K/AKT/glycogen synthase kinase-3
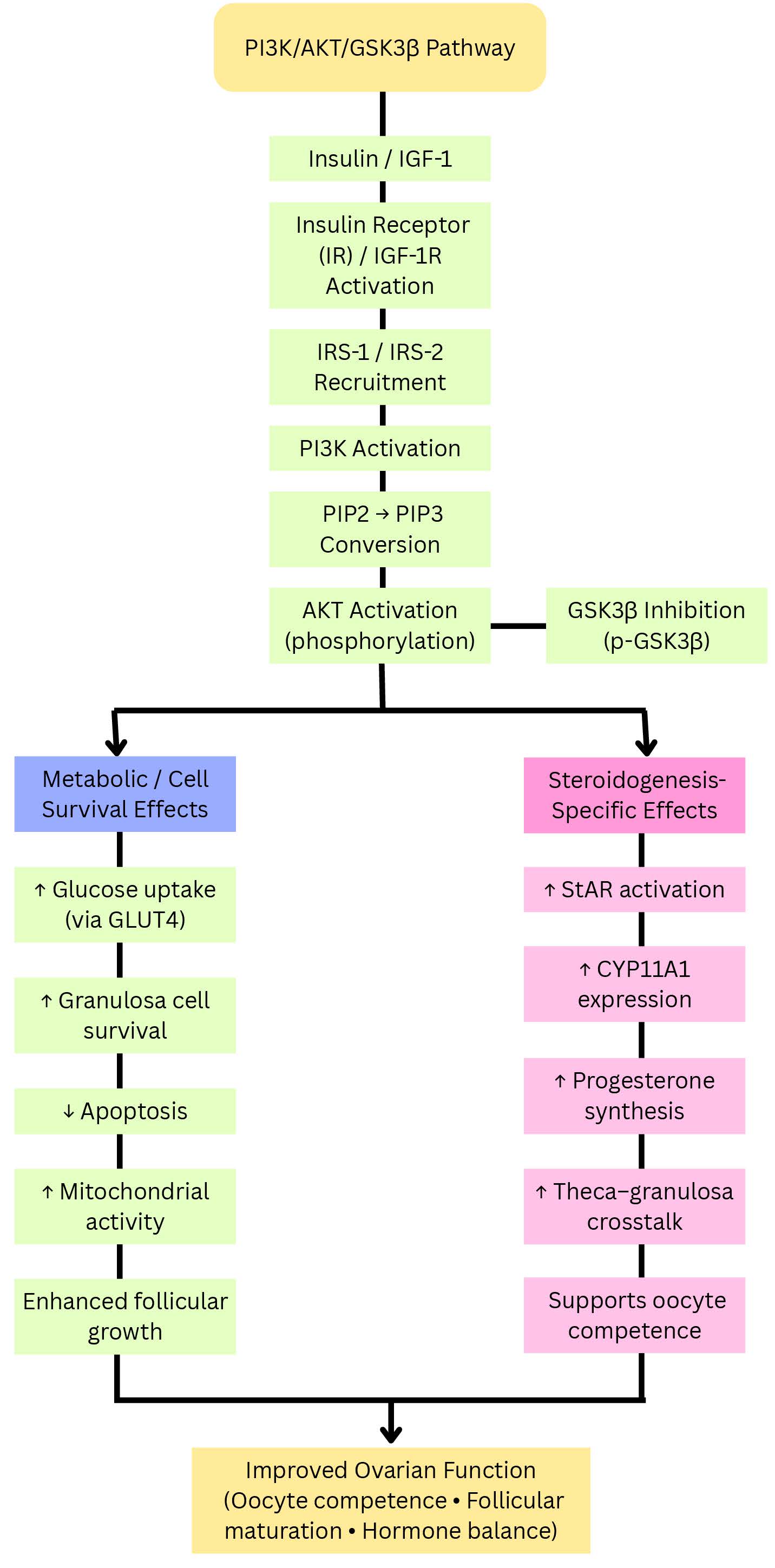 Fig. 5.
Fig. 5.
Split-branch representation of the PI3K/AKT/GSK3
AKT activation promotes estradiol synthesis and follicular maturation by
increasing the expression and activity of key enzymes such as CYP19A1 and StAR.
Additionally, the interaction between AKT and the transcription factor
transcription factor Forkhead Box O1 (FOXO1) regulates granulosa cell apoptosis
and guarantees follicular selection. Hyperinsulinemia and AKT hyperactivation
stimulate cytochrome P450 17A1 (CYP17A1) and HSD3
The MAPK and mTOR pathways act as metabolic sensors that link ovarian folliculogenesis to growth factor signaling, energy availability, and nutritional status (Fig. 6).
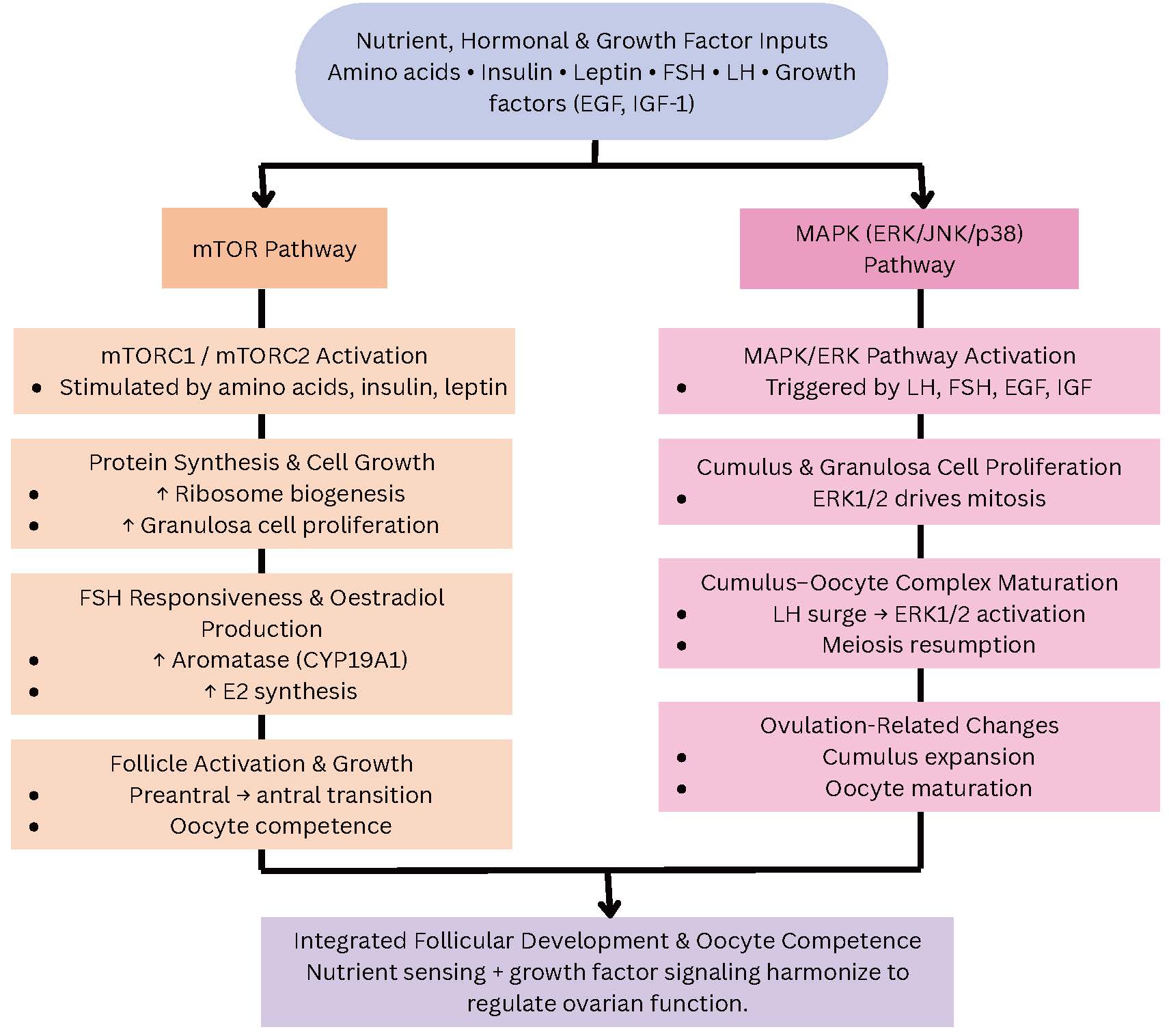 Fig. 6.
Fig. 6.
Parallel representation of the mTOR and MAPK pathways as nutrient- and hormone-sensitive regulators of follicular development. Amino acids, insulin, leptin, and gonadotropins activate mTORC1/2 and MAPK/ERK cascades in ovarian granulosa and cumulus cells. mTOR signaling enhances protein synthesis, cellular proliferation, aromatase expression, and follicle growth, whereas the MAPK pathway drives LH-induced cumulus expansion, meiosis resumption, and oocyte maturation. Together, these metabolic and growth factor-responsive pathways coordinate ovarian folliculogenesis, and their dysregulation contributes to follicle arrest and subfertility. Arrows indicate the direction of interaction or regulatory influence between components and do not represent quantitative effects. Abbreviations: EGF, epidermal growth factor; mTORC, mammalian target of rapamycin complex; ERK, extracellular signal-regulated kinase; E2, estradiol.
In response to amino acids, insulin, leptin, and mTOR, a serine/threonine kinase complex (mTORC1 and mTORC2), regulates protein synthesis, autophagy, and cell division. mTOR signaling stimulates the production of estradiol, FSH responsiveness, and cellular proliferation in ovarian granulosa cells. Its role in follicular activation and oocyte competence is supported by experimental inhibition, which decreases aromatase expression and hinders the preantral-to-antral follicle transition [18, 19].
In parallel, the MAPK/extracellular signalregulated kinase (ERK) pathway transmits growth factor and gonadotropin signals that regulate cumulus cell proliferation and oocyte maturation induced by the LH surge. Meiosis and cumulus-oocyte communication, two essential ovulation processes, are triggered by the LH-activated ERK1/2 cascade. mTOR and MAPK are also active in hypothalamic neurons, where they integrate GnRH secretion with nutrient-derived cues and link central reproductive regulation to peripheral metabolism. Dysregulation of these pathways, which may result from endocrine disruptors, obesity, or nutritional deprivation , can lead to follicular arrest and subfertility [20, 21].
Now known to play a significant role in ovarian dysfunction and anovulation, the
NF-
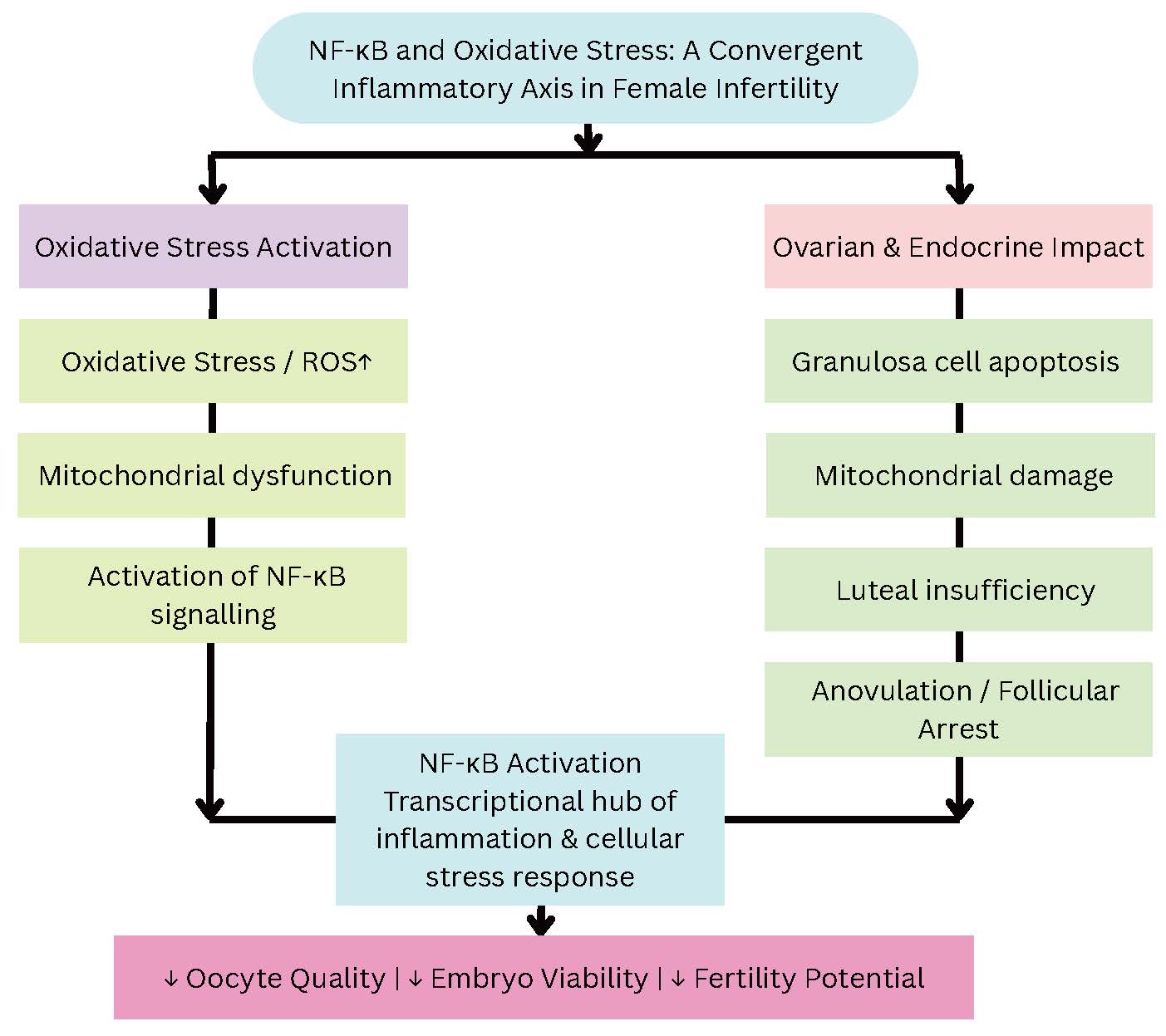 Fig. 7.
Fig. 7.
Integrative model illustrating how oxidative stress and chronic
activation of NF-
Oxidative stress suppresses GnRH release and modifies LH/FSH secretion by
interacting with NF-
AMPK, the primary cellular energy sensor in ovarian tissue, responds to alterations in the cellular adenosine monophosphate/adenosine triphosphate (AMP/ATP) ratio and metabolic cues by regulating follicle activation, granulosa cell proliferation, oocyte maturation, and autophagy. In the ovary, AMPK activation promotes metabolic adaptation in granulosa and cumulus cells, inhibits mTOR-driven follicle activation under low-energy conditions, and maintains oocyte quality by regulating mitochondrial and glucose metabolism. Experimental studies show that pharmacologic or genetic modulation of AMPK changes primordial follicle activation, follicular growth dynamics, and steroidogenic output. These findings suggest that reproductive disorders marked by decreased oocyte competence and impaired folliculogenesis are linked to dysregulated AMPK signaling (Fig. 8) [24].
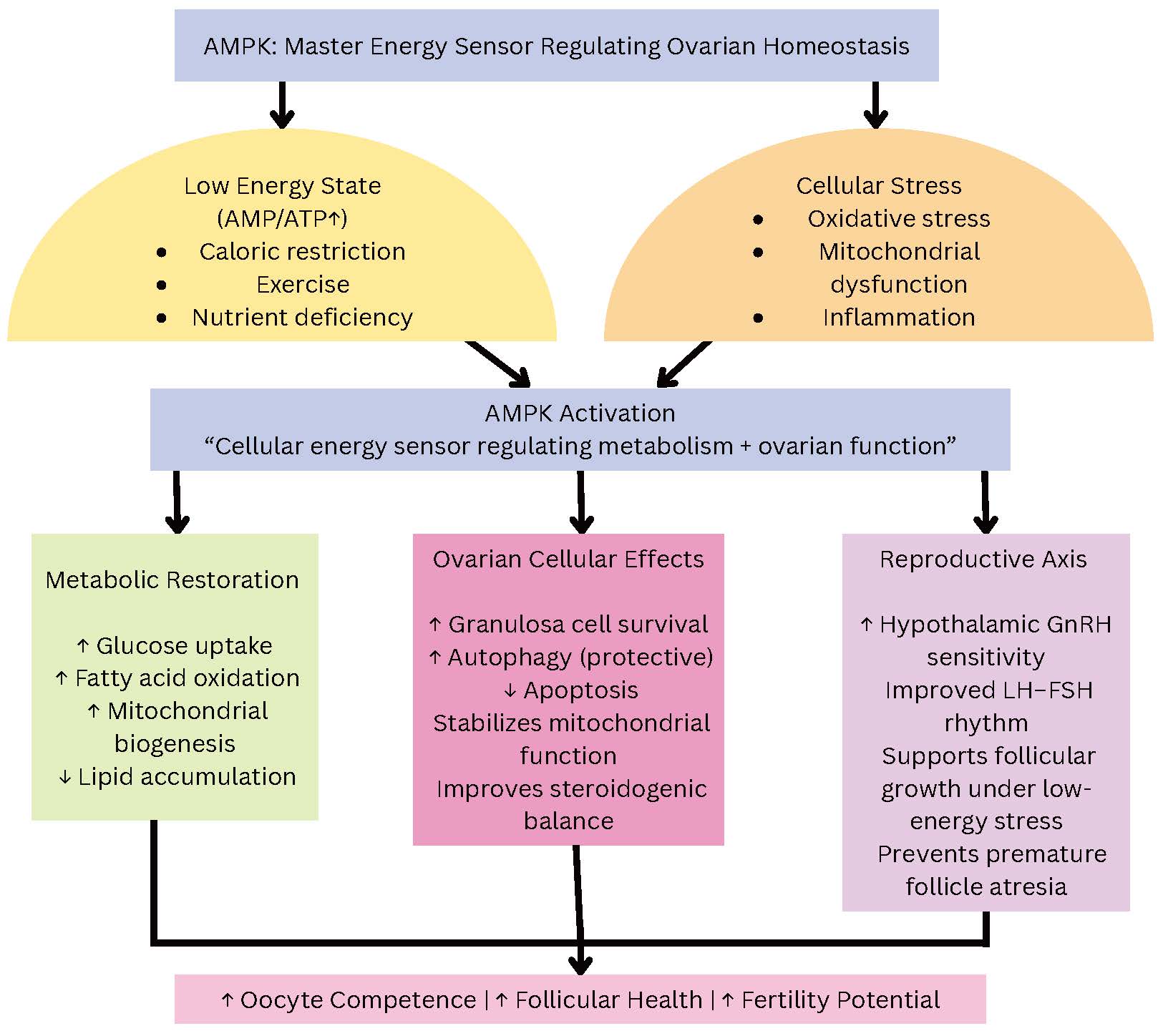 Fig. 8.
Fig. 8.
AMPK functions as a central metabolic sensor that integrates energy status, oxidative stress, and nutrient availability to maintain ovarian homeostasis. Low cellular ATP, mitochondrial stress, or inflammation activate AMPK, which subsequently restores metabolic balance through increased glucose uptake, fatty acid oxidation, and mitochondrial biogenesis. In ovarian cells, AMPK supports granulosa cell survival, regulates autophagy, prevents apoptosis, and maintains steroidogenic equilibrium. AMPK also interacts with hypothalamic-pituitary signaling to support GnRH and gonadotropin rhythms. Together, these actions preserve follicular development, oocyte competence, and fertility outcomes. Arrows indicate the direction of interaction or regulatory influence between components and do not represent quantitative effects.
The paracrine and autocrine signaling network formed by members of the
transforming growth factor-
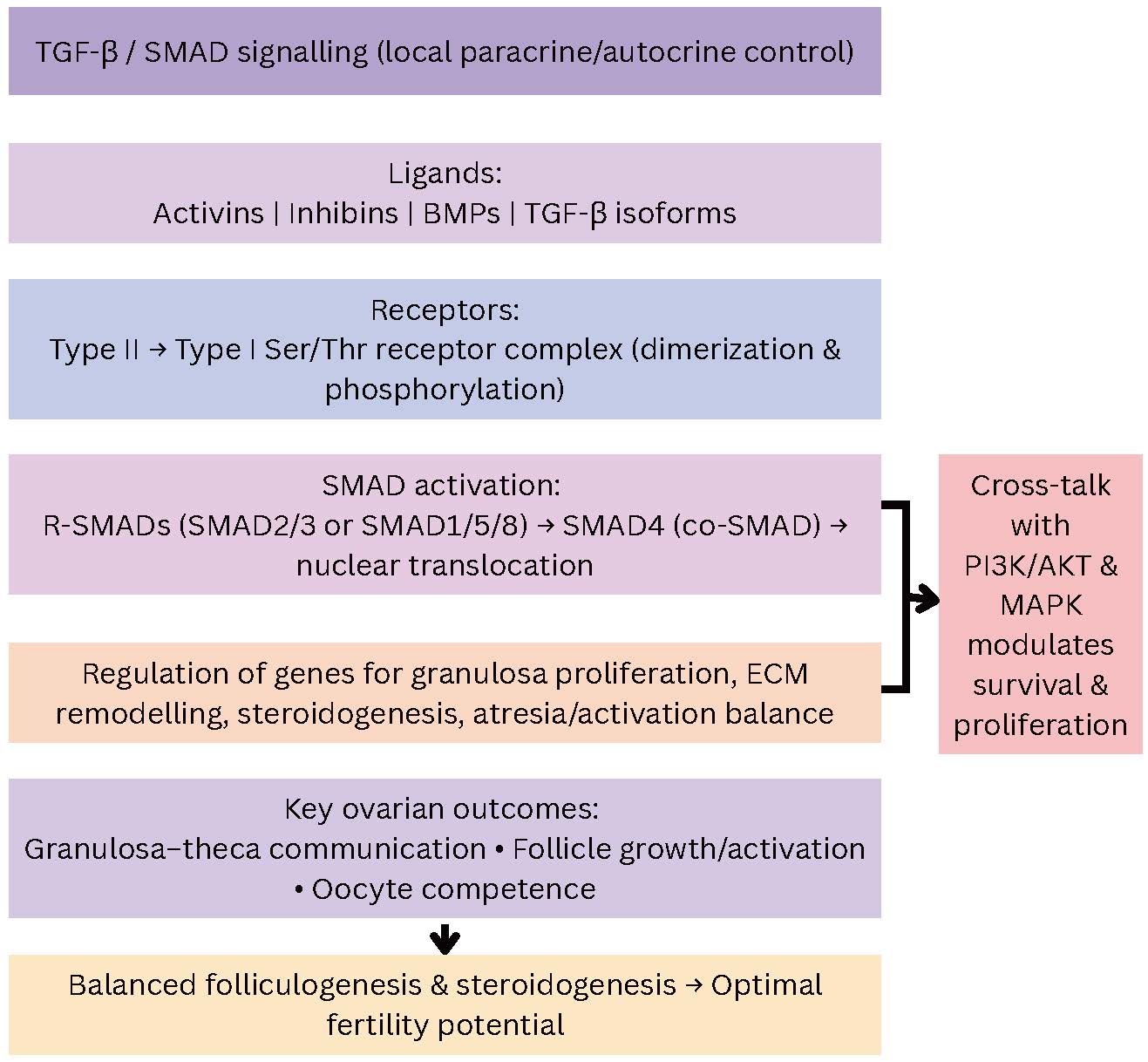 Fig. 9.
Fig. 9.
Simplified vertical representation of TGF-
An increasing number of research suggests that female fertility is associated with dietary intake. In this context, there is increasing interest in lifestyle factors, including diet and exercise, as well as psychological stress, socioeconomic factors, body mass index (BMI), smoking, alcohol, caffeine, and psychoactive substances. In infertility associated with endometriosis and ovulation difficulties, lifestyle factors, including calorie intake and diet composition in terms of vitamins, protein, lipids, carbs, and mineral content, appear to be particularly important (Fig. 10) [26, 27].
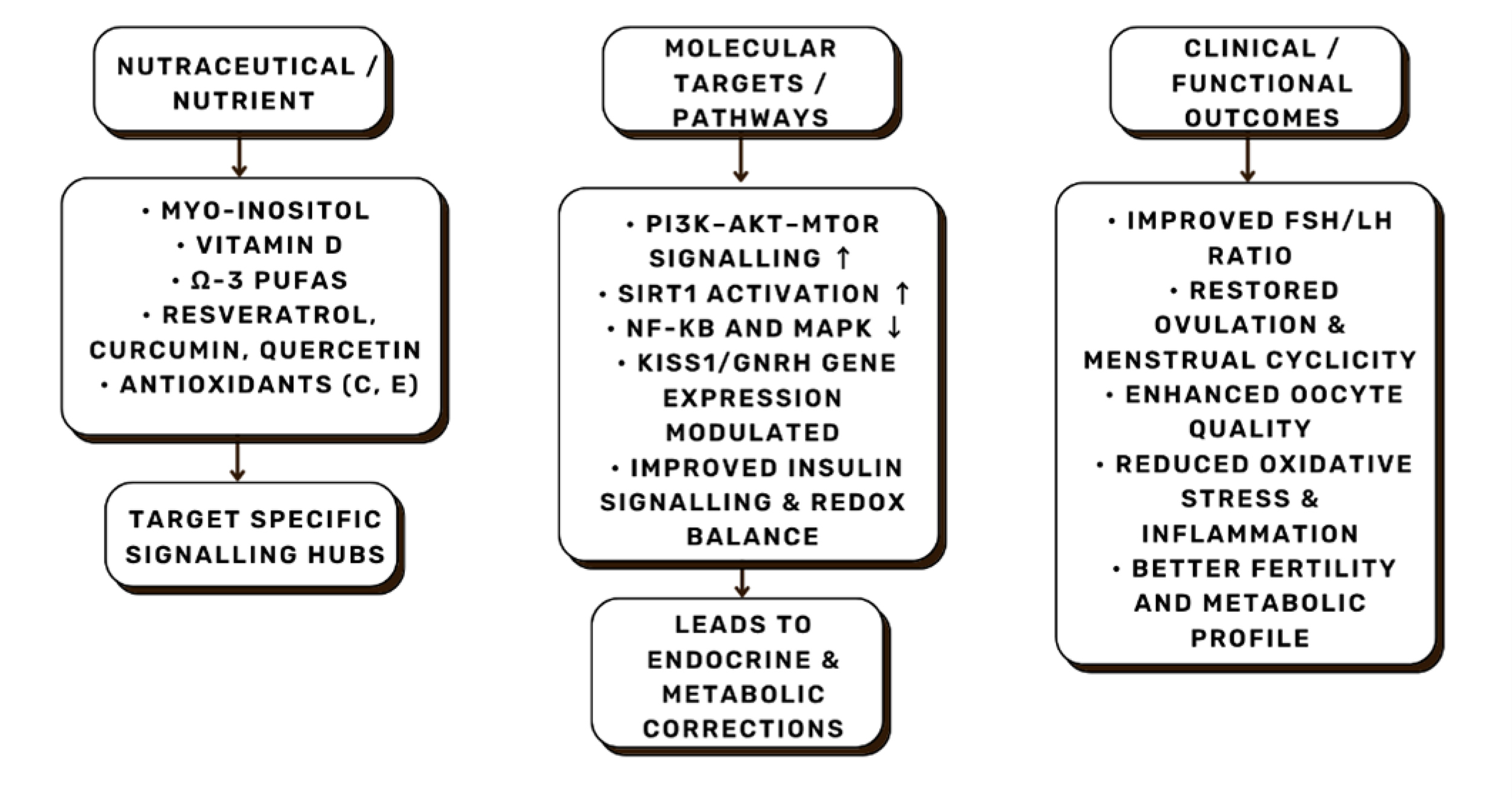 Fig. 10.
Fig. 10.
Representative nutrient–molecular–clinical outcome pathways.
Specific nutraceuticals modulate intracellular signaling hubs such as
PI3K/AKT/mTOR, SIRT1 (Sirtuin 1), and NF-
Diet and its nutritional components may have an impact on female fertility and ovulation through effects on metabolic pathways, endocrine profiles, and carbohydrate metabolism. Insulin resistance, the primary mechanism connecting anovulatory infertility to diet, is considered particularly significant in the development of ovulation disorders [28]. Oxidative stress and chronic inflammation are additional significant factors that raise the risk of ovulatory dysfunction [26]. Women with elevated inflammatory markers during their menstrual cycle are more likely to experience an anovulatory cycle, likely due to increased oxidative stress in the ovary [29].
The Nurses’ Health Study II (NHS II), a prospective cohort study involving 17,544 infertile women, found that diet and nutrient intake significantly impacted fertility and decreased the risk of ovulatory disorders. Compared with women in the lowest percentile, those in the highest quintile of adherence to a fertility-supporting diet had a 66% decreased chance of anovulatory infertility. Based on these findings, a “fertility diet” plan was proposed. It was characterized by the presence of high-fat dairy products, low-glycemic-index carbohydrates, high iron content, and a decrease in trans fatty acid consumption accompanied by an increase in monounsaturated fatty acid supply [28, 30].
Research in humans and animal models supports the concept that an appropriate diet enhances fertility. The Mediterranean diet (MD) in particular, has been extensively studied in this context [31, 32, 33, 34, 35]. Furthermore, numerous studies have also examined the relationship between the risk of infertility and intake of specific macronutrients or micronutrients, such as proteins, fats, carbohydrates, vitamins, and minerals like vitamin B12, vitamin D, folates, zinc, and omega-3 fatty acids.
The intricate interactions between metabolic, inflammatory, and neuroendocrine signals that control the HPO axis are influenced by nutrition, which is a major and modifiable factor in female reproductive health. Both macronutrients (proteins, fats, and carbohydrates) and micronutrients (vitamins, minerals, and trace elements) are essential for ovulation, steroidogenesis, and oocyte competence in women of reproductive age (Table 1, Ref. [36, 37, 38, 39, 40, 41, 42]). Dietary patterns associated with irregular menstruation, ovulatory dysfunction, and decreased fertility include high consumption of refined carbohydrates, trans fats, excess energy, and micronutrient deficiencies. In contrast, diets high in fiber, plant-based proteins, vitamins, minerals, and healthy fats, particularly omega-3 PUFAs, have positive correlations with reproductive outcomes [43, 44].
| Nutrient/Nutraceutical | Key molecular targets/pathways | Mechanistic action |
| Myo-/D-chiro-inositol | PI3K → AKT → GLUT4/CYP19A1 | Improves insulin signaling, enhances aromatase, reduces androgens [36, 37]. |
| Vitamin D | VDR, CYP19A1, IRS-1, GnRH | Regulates steroidogenesis, AMH, and cytokine balance [38]. |
| Omega-3 PUFAs | NF-κB, PPAR- |
Reduces inflammation, restores GnRH/kisspeptin signaling [39]. |
| NAC | NF-κB, Glutathione (GSH) axis | Antioxidant, insulin-sensitizer [40]. |
| CoQ10 | Mitochondrial ETC Complex I–III | Enhances ATP, oocyte maturation [40]. |
| Melatonin | MT1/MT2 receptors, SIRT3 | Antioxidant, circadian and steroidogenic regulation [40]. |
| Zinc | Metallothionein, Meiotic enzymes | Chromatin stability, oocyte meiosis [41]. |
| Folate/B12 | DNA methyltransferase, MTHFR | Epigenetic regulation of GnRH/steroidogenesis [41]. |
| Polyphenols (resveratrol, curcumin) | SIRT1, AMPK, Nrf2 | Anti-inflammatory, energy-sensing modulation [42]. |
VDR,vitamin D receptor; NAC, N-acetylcysteine; CoQ10, coenzyme Q10; MTHFR, methylenetetrahydrofolate reductase; AMH, anti-Müllerian hormone; PUFAs, polyunsaturated fatty acids.
Building on the dietary basis, a “nutraceutical” is defined as a food or component of a food that has health-promoting or medical effects, such as preventing or treating disease. These products are frequently supplied in concentrated, formulated form rather than whole foods alone. By enabling modulation of specific pathways linked to reproductive dysfunction, nutraceuticals act as a link between targeted molecular interventions and general nutrition. Nutraceuticals address the key disruptions of the neuroendocrine-ovarian axis, including insulin resistance, oxidative stress, inflammation, and altered steroidogenesis, in the context of female infertility. According to a comprehensive analysis of dietary supplements, common nutraceuticals like inositols, vitamin D, omega-3 fatty acids, and antioxidants, are frequently used to support fertility [45, 46].
A balanced macronutrient composition and adequate micronutrient intake provide the foundation for optimal neuroendocrine signaling. Impaired ovarian function, irregular menstrual cycles, and an elevated risk of fertility issues are associated with deficiencies in vitamins (D, B-complex), minerals (iron, selenium, and zinc), and fatty acid intake. Diets rich in vegetables, whole grains, lean meats, and omega-3 PUFAs have been linked to improved in vitro fertilization (IVF) outcomes, such as higher oocyte quality and embryo yields, confirming the concept that nutrition affects the ovarian microenvironment and hypothalamic-pituitary signaling. Mechanistically, nutrients affect insulin sensitivity, inflammatory cytokines, oxidative stress pathways, and adipokines (adiponectin, leptin), all of which influence the release of gonadotropin, the secretion of GnRH, and ovarian steroidogenesis [47, 48].
4.3.2.1 Myo-inositol/D-chiro-inositol
The stereoisomers of inositol, myo-inositol (MI) and D-chiro-inositol (DCI), function as intracellular second messengers for insulin and gonadotropin (FSH/LH) signaling. MI improves insulin sensitivity, lowers hyperinsulinemia, and attenuates ovarian androgen excess by modulating the PI3K/AKT pathway and promoting glucose transporter 4 (GLUT-4) translocation in granulosa and theca cells. In insulin signaling, MI functions as a secondary messenger by triggering the PI3K/AKT pathway, which phosphorylates and activates GLUT-4 vesicle transporters, leading to their translocation to the cell membrane. As a result, granulosa and theca cells exhibit increased glucose uptake, which reduces intracellular glucose accumulation and insulin resistance. Compensatory hyperinsulinemia is decreased, and ovarian androgen synthesis is subsequently reduced, as insulin amplifies LH-induced androgen production in theca cells. MI supplementation improves oocyte quality, increases ovulation rates, and lowers the requirement for exogenous FSH needed for IVF procedures. MI directly connects metabolic modulation to HPO axis correction at the neuroendocrine level by promoting follicular responsiveness to gonadotropins and restoring the FSH/LH ratio [36, 37].
4.3.2.2 Vitamin D
In addition to maintaining calcium homeostasis, the active form of vitamin D
binds to the vitamin D receptor (VDR), which is expressed in the pituitary,
ovarian, and hypothalamic tissues. It decreases the expression of
pro-inflammatory cytokines (IL-6, TNF-
4.3.2.3 Omega-3 PUFAs
The primary dietary omega-3 PUFAs, eicosapentaenoic acid (EPA) and
docosahexaenoic acid (DHA), inhibit the NF-
4.3.2.4 Antioxidants and Mitochondrial Protectors (N-acetylcysteine, CoQ10, Melatonin, Vitamins C/E)
Oxidative stress and mitochondrial dysfunction are major threats to oocyte
quality, as they hinder meiotic spindle formation, increase DNA damage, and
diminish embryo competence. Vitamins C and E restore antioxidant capacity;
melatonin acts as a direct free radical scavenger and modulates circadian
steroidogenic rhythms; coenzyme Q10 (CoQ10) increases mitochondrial respiratory
chain efficiency; N-acetylcysteine (NAC) replenishes glutathione and inhibits
NF-
4.3.2.5 Micronutrients (Zinc, Folate/B-Vitamins, Iron, Selenium)
Chromatin stability, DNA methylation, oocyte maturation, and embryo development are maintained by vital trace elements and vitamins. Iron and selenium regulate oxidative status and interactions between the thyroid and reproductive axis; zinc is essential for meiotic progression and antioxidative enzyme activity; folate and B vitamins regulate one-carbon metabolism and epigenetic programming of genes involved in GnRH regulation and steroidogenesis. A lower ovarian reserve, a higher risk of miscarriage, and poor embryo quality are associated with suboptimal micronutrient status [41].
4.3.2.6 Polyphenols and Botanical Adaptogens
Polyphenols with pleiotropic effects on metabolic and neuroendocrine pathways
include quercetin, curcumin, and resveratrol. These compounds exert dual
regulation of metabolic homeostasis and hypothalamic-pituitary signaling by
activating SIRT1 and Nrf2, downregulating mTOR and NF-
4.3.3.1 Mediterranean Diet (MD)
Improved fertility outcomes have most consistently been linked to the MD,
which is rich in fruits, vegetables, legumes, whole grains, nuts,
extra-virgin olive oil, moderate amounts of fish, and low amounts of red meat.
Higher adherence to the MD is associated with higher rates of clinical pregnancy
and embryo implantation in women undergoing assisted reproductive technologies
(ART), especially in younger cohorts. Mechanistically, its high concentration of
antioxidants, polyphenols, and omega-3 fatty acids lowers oxidative stress,
improves insulin sensitivity, and modulates inflammatory cytokines
(TNF-
4.3.3.2 Low-Glycemic (GI) and Dietary Approaches to Stop Hypertension (DASH) Diets
In women with insulin resistance, low-GI diets that prioritize whole grains, legumes, and minimal amounts of refined sugars improve insulin dynamics and restore ovulatory cycles. After 12 weeks of a low-GI diet, randomized trials in PCOS patients show improved LH/FSH ratios, decreased fasting insulin, and increased ovulation frequency. This is explained by downstream modulation of insulin receptor/IRS-1/PI3K-AKT signaling, which promotes follicular growth and decreases the production of androgen by theca cells. Similarly, women with reproductive endocrine disorders who follow the Dietary Approaches to Stop Hypertension (DASH) diet, which is low in saturated fats and rich in fruits, vegetables, and low-fat dairy, have shown improved hormonal profiles and decreased inflammatory biomarkers [52, 53].
4.3.3.3 Plant-Based and Anti-Inflammatory Diets
Recent studies also suggest that plant-forward or anti-inflammatory diets, which
are high in antioxidants, fiber, and phytonutrients, can improve fertility. By
fostering microbial diversity and gut-brain-ovary axis communication, these diets
reduce oxidative load and systemic inflammation, two factors that significantly
impair hypothalamic GnRH pulsatility. Foods rich in polyphenols, like soy,
berries, and green tea, are especially noteworthy for their ability to modulate
the SIRT1, Nrf2, and NF-
Recent biomarker panels for evaluating female reproduction include metabolic signals and neuropeptides in addition to conventional hormones. AMH remains the most widely used indicator of ovarian reserve and response in assisted reproduction; however, its interpretation depends on several factors, including age, assay, and clinical context [55]. Recent research has indicated that serum kisspeptin, a direct regulator of GnRH neurons, may serve as a useful marker for neuroendocrine evaluation, reflecting hypothalamic-pituitary integrity and early pregnancy outcomes [56]. An elevated LH:FSH ratio remains a useful indicator of neuroendocrine imbalance in PCOS, despite interindividual variability. Adipokines, such as leptin, reflect energy status and are consistently elevated in PCOS, connecting metabolic phenotype with ovarian dysfunction [57]. Lastly, circulating nutrient biomarkers, such as omega-3 fatty acid profile and 25-hydroxyvitamin D (25-OH D), offer objective assessments of nutritional status that correlate with ovarian function and ART outcomes. These measures can also be used in conjunction with endocrine panels [58].
Because vitamin D status influences ovarian steroidogenesis, AMH, and implantation biology, measurement of serum 25-OH D is a useful diagnostic tool in infertility clinics. Although conclusive RCT evidence is still limited, both observational and interventional studies have reported associations between higher 25-OH D levels and improved IVF/ART parameters. An objective measure of anti-inflammatory lipid status can be obtained from the omega-3 index (erythrocyte EPA+DHA%) or serum long-chain omega-3 levels. In certain ART cohorts, higher maternal omega-3 status has been linked to lower pregnancy loss and improved live birth probabilities. Incorporation of these nutrient biomarkers into preconception evaluations provides a quantifiable target for corrective nutritional interventions [58, 59].
Patient stratification and meticulously developed nutraceutical formulations are necessary to translate single-agent evidence into clinical practice. By customizing interventions to metabolic and inflammatory profiles (e.g., insulin-resistant vs. inflammatory phenotypes), precision nutrition approaches, which combine diet, targeted supplements, and individual phenotype or genotype data, have shown early promise in improving fertility endpoints. While individualized dietary plans and customized nutraceutical combinations represent the obvious next step in improving reproductive outcomes, narrative and systematic reviews emphasize the need for carefully controlled trials to determine the best formulations, dosages, and treatment windows [60].
AI and ML algorithms, as well as molecular mapping approaches, including pathway
and PPI analyses, are quickly emerging as screening and prioritization tools in
reproductive medicine. While AI models trained on clinical, biochemical, and
imaging data have shown early success in risk stratification, embryo selection,
and outcome prediction in ART, network-based approaches can identify convergent
molecular hubs (e.g., PI3K/AKT, mTOR, NF-
Although several nutrients show promising benefits for reproductive function, the available evidence is limited by the scarcity of large, multicenter clinical trials. Variability in population characteristics, including ethnicity, age, and baseline metabolic status, further influence outcomes and limits generalizability. Many findings derive from small human cohorts, animal models, or short-term interventions, which restrict the generalizability of nutrient–neuroendocrine interactions in female fertility. Additionally, multi-omics datasets used for pathway enrichment and molecular network interpretations remain incomplete, and gene–nutrient associations may vary across populations, age groups, and infertility etiologies. As high-throughput technologies continue to evolve, larger, well-controlled clinical and transcriptomic studies are required to validate emerging mechanistic insights and strengthen the translational relevance of the pathways discussed in this review.
Female infertility arises from a complex network involving ovarian function,
metabolic homeostasis, nutritional status, and neuroendocrine signaling from the
brain. The “brain-to-ovary” continuum emphasizes that restoring fertility
requires the modulation of the neuroendocrine and metabolic axes that control
reproductive organs, rather than focusing solely on the organs themselves. This
review integrates mechanistic and translational evidence supporting the ability
of nutrients and nutraceuticals to alter reproductive physiology at both the
central and peripheral levels. Key intracellular pathways that regulate ovarian
steroidogenesis and oocyte competence, including PI3K/AKT/mTOR, NF-
MN conceptualized the study, conducted the literature review, performed data curation and synthesis, and drafted the manuscript. BJ and RN supervised the study, contributed to the organization and critical revision of the review, and provided expert guidance. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
We thank all colleagues and mentors who supported the development of this manuscript. We also extend our appreciation to the peer reviewers for their constructive insights that strengthened the quality of this work.
This research received no external funding.
The authors declare no conflict of interest.
During the preparation of this manuscript, the authors used ChatGPT (OpenAI, GPT-5.3) for grammar checking and language polishing. After using this tool, the authors reviewed and edited the content manually and take full responsibility for the final text.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.