

 , Fangyuan Luo 2, Jinpeng Wang 3,*
, Fangyuan Luo 2, Jinpeng Wang 3,*
1 Department of Gynecology and Obstetrics, Deyang People’s Hospital, 618099 Deyang, Sichuan, China
2 Department of Obstetrics and Gynecology, Sichuan Provincial People’s Hospital, University of Electronic Science and Technology of China, 610091 Chengdu, Sichuan, China
3 Traditional Chinese Medicine Department, Reproductive & Women-Children Hospital, Chengdu University of Traditional Chinese Medicine, 610041 Chengdu, Sichuan, China
Abstract
This review systematically examines the multifaceted roles of Human leukocyte antigen F (HLA-F) in the tumor and maternal-fetal interface microenvironments.
A comprehensive literature search was conducted using PubMed, Medical Literature Analysis and Retrieval System Online (MEDLINE), and Embase databases, with a focus on studies published within the past five years. Keywords included “HLA-F”, “tumor microenvironment (TME)”, “maternal-fetal interface”, “immune tolerance”, and “preeclampsia”. To our knowledge, this study represents the first integrative analysis to delineate the context-dependent immunoregulatory functions of HLA-F.
HLA-F exhibits dual and contrasting functions. In cancer, it facilitates immune evasion and correlates with poor prognosis via interactions with inhibitory receptors. At the maternal-fetal interface, it promotes immune tolerance, which is essential for pregnancy maintenance.
HLA-F serves as a pivotal immunoregulatory molecule, with significant implications for cancer immunotherapy and reproductive medicine.
Keywords
- HLA-F
- tumor microenvironment
- maternal-fetal interface
- immune tolerance
- preeclampsia
The human leukocyte antigen (HLA), located on chromosome 6, is a critical component of the human immune system [1]. This polymorphic gene family encodes cell surface proteins essential for antigen presentation and immune regulation [1, 2]. HLA molecules are broadly classified into class I (HLA-A, -B, -C) and class II (HLA-DP, -DQ, -DR). Classical class I molecules (HLA-A, -B, -C) present endogenous peptides to CD8+ T cells, eliciting cytotoxic responses against infected or malignant cells [2]. Class II molecules present exogenous antigens to CD4+ T helper cells, orchestrating broader immune activation [2]. In contrast, non-classical class Ib molecules, including HLA-E, HLA-F, and HLA-G, exhibit limited polymorphism and perform specialized immunoregulatory functions, typically modulating immune responses in specific physiological and pathological contexts [3, 4].
HLA-F, a member of the non-classical HLA class Ib family initially described as HLA-5.4 by Geraghty et al. [5] in 1989, has emerged as an important immunomodulatory molecule. Unlike classical HLA molecules, HLA-F can bind to both activating and inhibitory receptors expressed on immune cells as well as other receptor-expressing cells, such as V-domain immunoglobulin suppressor of T-cell activation (VISTA) on trophoblasts [6], positioning it as a critical regulator of immune tolerance and activation across diverse immune microenvironments. Its expression is highly restricted in healthy tissues but is frequently upregulated under specific circumstances, notably in cancer and at the maternal-fetal interface [7].
Recent studies have highlighted the dual roles of HLA-F in the tumor microenvironment (TME) and maternal-fetal interface [3, 8]. In various malignancies, including melanoma and breast and ovarian cancers, HLA-F overexpression is often associated with immune evasion, poor clinical outcomes, and therapeutic resistance [7, 9]. HLA-F interacts with receptors such as killer cell immunoglobulin like receptor, three Ig domains and short cytoplasmic tail 1 (KIR3DS1) on natural killer (NK) cells and immunoglobulin-like transcript 2/4 (ILT2/4) on other immune cells, potentially suppressing NK cell cytotoxicity and contributing to an immunosuppressive TME that facilitates tumor progression [8]. Conversely, at the maternal-fetal interface, controlled expression of HLA-F is considered essential for establishing immune tolerance toward the semi-allogeneic fetus [3]. HLA-F is widely expressed on syncytiotrophoblast, extravillous trophoblasts (EVTs), and various maternal-fetal immune cells, where it likely supports successful placentation and pregnancy maintenance [10]. Rather than representing opposite functions, HLA-F exerts a shared immunoregulatory program in both contexts. This program is physiologically beneficial at the maternal-fetal interface, where immune tolerance is required for fetal survival, but becomes pathologically disadvantageous for the host in cancer, where the same mechanisms are co-opted to facilitate tumor immune escape.
HLA-F has emerged as a multifaceted immunoregulatory molecule whose study provides important insights into immune tolerance, cancer biology, and reproductive immunology. Notably, the tumor microenvironment shares striking conceptual and functional parallels with the maternal-fetal interface, particularly with respect to immune adaptation, tissue remodeling, and tolerance induction. Within these distinct yet convergent biological contexts, HLA-F operates through diverse molecular conformations and receptor interactions to shape local immune responses. In this review, we synthesize current knowledge on the involvement of HLA-F in tumorigenesis, female reproductive physiology, and pregnancy-related disorders, and discuss how emerging mechanistic insights may inform future therapeutic strategies. An overview of HLA-F molecular forms, receptor engagements, and their context-dependent outcomes in cancer and pregnancy is provided in Fig. 1.
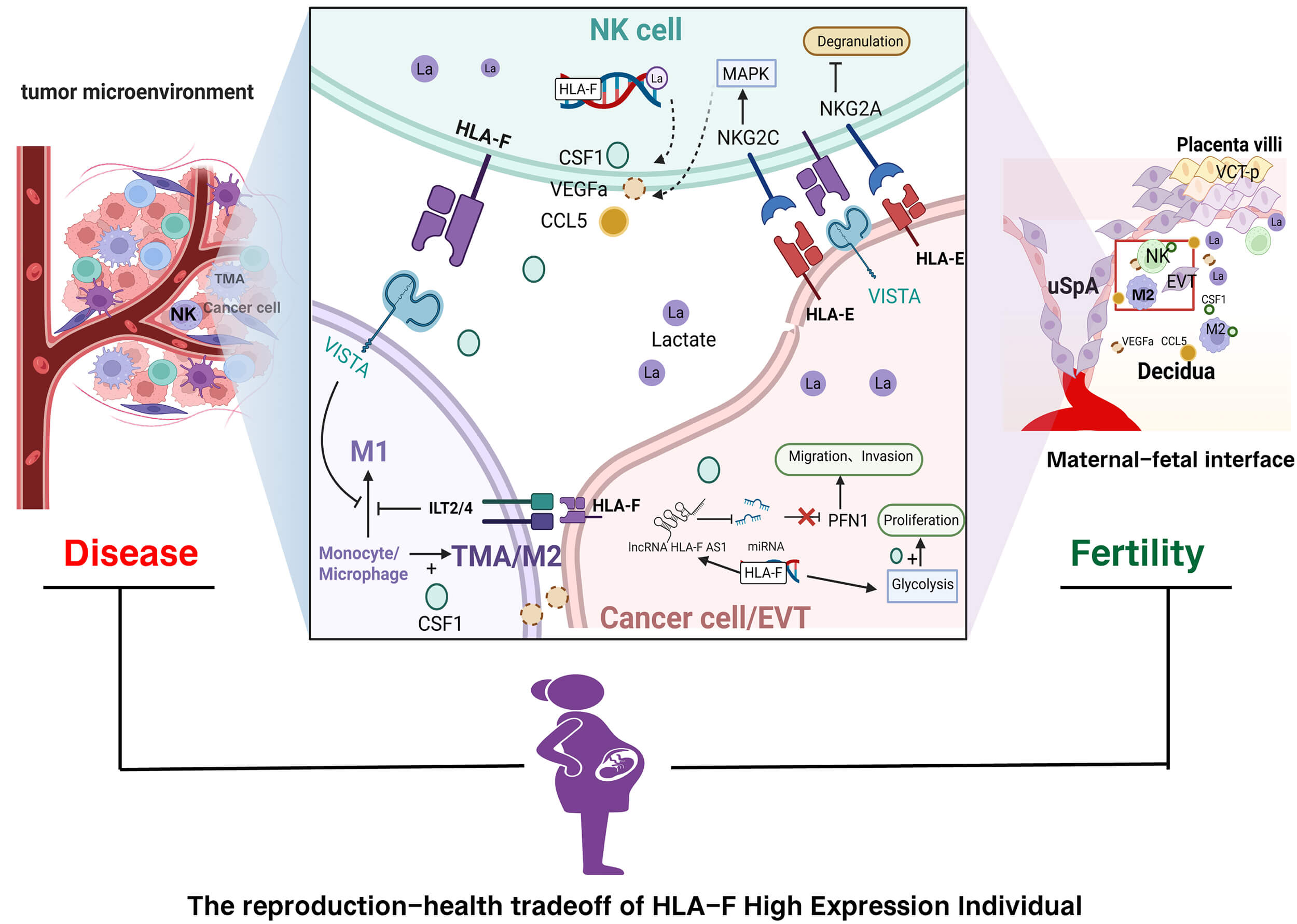 Fig. 1.
Fig. 1.
HLA-F conformers and context-specific immunoregulation. HLA-F
exists in distinct molecular conformations, including
Although 45 HLA-F haplotypes (alleles) have been identified, they encode only
six protein variants, indicating that the HLA-F gene has remained highly
conserved during human evolution [11]. HLA-F mRNA is widely expressed in multiple
tissues and organs, with particularly high expression levels in the spleen,
lungs, and digestive tract tissues. At the cellular level, HLA-F shows the
highest expression in activated immune cells, followed by relatively uniform,
high expression in epithelial cells, endothelial cells, adipocytes, and glial
cells (https://www.proteinatlas.org). The classical function of HLA-F involves
forming a transmembrane
Apart from antigen presentation, HLA-F can exist on the cell surface as
peptide-free heavy chains (open conformers, OCs), interacting with immune
receptors on immune cells. HLA-F open conformers can form dimeric transmembrane
proteins that interact with inhibitory receptors killer cell immunoglobulin-like
receptor, three Ig domains and long cytoplasmic tail 1/killer cell
immunoglobulin-like receptor, three Ig domains and long cytoplasmic tail 2
(KIR3DL1/KIR3DL2) and activating receptors killer cell immunoglobulin-like
receptor, three Ig domains and short cytoplasmic tail 1/killer cell
immunoglobulin-like receptor, two Ig domains and short cytoplasmic tail 4
(KIR3DS1/KIR2DS4) on NK cells [16, 17]. Since the binding sites for ILT2
(leukocyte immunoglobulin-like receptor subfamily B member 1 [LILRB1]) and ILT4
(leukocyte immunoglobulin-like receptor subfamily B member 2 [LILRB2]) are
located outside the peptide-binding groove, HLA-F OCs, similarly to
Furthermore, HLA-F open conformers can localize to the nucleus, cytoplasm, and extracellular matrix, exhibiting diverse protein-level functions [10, 19, 20]. Proteomic analysis of HLA-F allele-specific peptides identified glycolytic enzymes, including pyruvate kinase M (PKM) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH), among peptides restricted by HLA-F*01:01 isolated from LCL721.221 cells [21], suggesting an association between HLA-F and glycolytic pathway enzyme activities. A study in glioma cells revealed that HLA-F interacts with hexokinase 2 (HK2), extending its half-life, thereby enhancing enzymatic activity, glycolysis, and cell proliferation in glioma [22]. This observation highlights HLA-F’s role in promoting glycolysis through interactions with glycolytic enzymes. A recent study also showed that HLA-F binds the PKM promoter region, enhancing PKM2 expression and consequently increasing glycolysis and proliferation in trophoblasts [23]. This finding represents the first evidence of HLA-F functioning as a transcriptional regulator beyond its immunomodulatory role. Collectively, these studies illustrate that cells highly expressing HLA-F exhibit elevated glycolysis, consistent with previous single-cell sequencing findings at the maternal-fetal interface, demonstrating that the dNK1 subtype, characterized by higher HLA-F expression, exhibits increased glycolytic metabolism [24].
In addition to its protein-level roles, the long non-coding RNA of HLA-F (lncRNA HLA-F-AS1) regulates gene transcription through multiple pathways, including targeting miR-21-3p, thereby promoting tumor cell migration and invasion [25, 26, 27].
The TME comprises cells of both innate and adaptive immunity, as well as non-immune cells such as fibroblasts and endothelial cells [28]. Within the TME, HLA-F is expressed not only by tumor cells [7], but also by immune cells such as NK cells and tumor-associated macrophages (TAMs) [10]. Moreover, certain tumor cells themselves express the inhibitory HLA-F receptor VISTA [6], establishing bidirectional immune tolerance between tumor and immune cells. Numerous studies have reported that elevated expression of HLA-F or LncRNA HLA-F-AS1 correlates with poor prognosis in glioma [7, 22, 27, 29], breast cancer [30, 31], ovarian cancer [25], gastric cancer [32], lung cancer [9], and colorectal cancer [26]. Collectively, these findings indicate that HLA-F plays an essential role in immune escape by tumor cells, significantly promoting immune tolerance within the TME.
The mechanisms by which HLA-F participates in immune escape within the TME include enhanced glycolysis-mediated proliferation of tumor cells due to high HLA-F expression [33]. Additionally, the lactate-rich microenvironment (the Warburg effect) within the TME facilitates metabolic reprogramming of immune cells through post-translational modifications, thereby suppressing the cytotoxic activities of NK cells and TAMs, ultimately enabling immune evasion [28, 34]. Previous studies demonstrated that HLA-F promotes glycolysis in glioma and trophoblastic tumor cells [22, 23]. Although direct studies linking HLA-F with lactylation modifications in immune cells remain absent, it is plausible that elevated HLA-F expression in immune cells similarly enhances glycolysis and regulates immune functions via lactylation. Research indicates that inhibiting lactylation in NK cells increases their cytotoxicity against tumor cells [34], suggesting another potential mechanism by which HLA-F facilitates immune escape. Furthermore, LncRNA HLA-F-AS1 interacts with various microRNAs to regulate tumor cell biological behaviors [25, 26, 27, 30]. However, the predominant mechanism underlying immune evasion is tumor-expressed HLA-F directly binding inhibitory receptors (e.g., VISTA, ILT2/ILT4, KIR3DL1/KIR3DL2) on immune cells, thereby suppressing immune functions [28].
Among the various HLA-F receptors, VISTA is the most recently identified and
least studied [18]. As previously described, HLA-F binds to VISTA as a
heterodimer [18]. VISTA (also known as VISR) is a novel inhibitory immune
checkpoint receptor crucial to HLA-F-mediated immune tolerance within the TME
[35]. Like cytotoxic T-lymphocyte antigen-4 (CTLA-4) and programmed death 1 (PD-1)/programmed cell death ligand-1 (PD-L1), VISTA belongs to the B7 family. Despite
significant sequence homology with PD-L1, VISTA functions via distinct mechanisms
and may therefore enable combination therapies [36]. In peripheral and
circulating immune cells, VISTA is constitutively expressed on naïve T
cells, independently inhibiting T-cell activation to maintain T-cell quiescence
[37, 38] and promoting drug resistance against anti-tumor agents [39]. Within the
TME, VISTA is highly expressed in myeloid-derived suppressor cells (MDSCs),
particularly TAMs [28, 40]. Under hypoxic conditions, VISTA expression is
significantly elevated, reducing toll-like-receptor (TLR) signaling and cell migration [41].
Furthermore, VISTA reprograms myeloid populations, decreasing pro-inflammatory
cytokines (e.g., tumor necrosis factor-alpha [TNF-
Maternal-fetal immunity and the TME exhibit notable parallels in their immunoregulatory strategies, including the establishment of local immune tolerance, modulation of NK cell activity, and extensive vascular remodeling [16]. During pregnancy, extravillous trophoblasts mediate spiral artery remodeling to generate low-resistance vessels that support placental development [20], whereas in cancer, comparable vascular adaptations, such as vascular mimicry, facilitate tumor growth [32]. In both contexts, immune cells are functionally reprogrammed toward a tolerogenic phenotype, thereby limiting immune-mediated tissue damage [16, 32]. Within this shared immunological framework, non-classical HLA molecules, including HLA-F, are thought to contribute to immune adaptation in both physiological pregnancy and pathological tumor progression.
The maternal-fetal interface refers to the interaction zone between maternal decidual immune and stromal cells and fetal trophoblasts and immune cells. Some scholars propose that this interface consists of two distinct layers. The first interface involves interactions between fetal EVTs and maternal decidual immune cells. The second interface comprises interactions between syncytiotrophoblast on placental villi and maternal circulating immune cells, as well as the extended interface formed by trophoblast-derived debris, including single trophoblast cells, cell fragments, and microvesicles, interacting with maternal immune cells and endothelial cells in the circulation [46]. Both interfaces are present throughout gestation; however, during early pregnancy, immune interactions at the EVT-decidual interface play a dominant role in implantation and placentation, whereas contributions from the syncytiotrophoblast–circulating immune interface become increasingly prominent as placental development progresses [46]. Interactions between fetal and maternal cells at the interface begin at embryo implantation and persist throughout gestation [24, 47]. Because the human embryo contains paternal antigens, it represents a semi-allogeneic graft to the maternal immune system [48]. Furthermore, EVTs exhibit invasive behavior similar to tumor cells, penetrating the decidua and uterine spiral arteries. Thus, immune tolerance at the maternal-fetal interface shares many mechanistic features with immune escape in the TME.
Aside from post-ovulatory endometrial decidualization and decidual recruitment of immune-tolerant immune cell subsets, another key mechanism involves trophoblast expression of non-classical HLA class Ib molecules, similar to tumor cells that upregulate HLA-F and related molecules. During early pregnancy, EVTs, which are in direct contact with maternal immune cells, express low levels of classical, paternally derived HLA-C and primarily express low-polymorphic non-classical molecules HLA-G, HLA-F, and HLA-E [48]. Among these, HLA-G is markedly upregulated during the differentiation of villous cytotrophoblasts (VCTs) into EVTs, serving as a dominant ligand mediating EVT-immune cell communication. In early decidua, NK cells constitute the major immune population, whereas monocytes and macrophages increase progressively during mid- to late-pregnancy. HLA-G receptors are widely distributed on immune cells at the maternal-fetal interface. HLA-G can bind KIR2DL4 and LILRB1 on NK cells, suppressing their cytotoxic activity; engage ILT4 and ILT2 on macrophages and dendritic cells to inhibit dendritic cell maturation and reduce major histocompatibility complex class II (MHC-II) expression, thereby maintaining immune tolerance; and interact with TCR and ILT2 on CD8+ and CD4+ T cells to suppress cytotoxicity, induce CD8+ T-cell apoptosis, and inhibit CD4+ T-cell proliferation [49]. Soluble HLA-G may also bind HLA-E to form stable complexes that interact with cluster of differentiation 94/natural killer group 2 (CD94/NKG2) receptors, further inhibiting NK cell cytotoxicity [50, 51]. The EVT-specific expression pattern and potent immunosuppressive effects of HLA-G underscore its role as a key “terminal effector” or “frontline soldier” in establishing and maintaining maternal-fetal immune tolerance.
HLA-G is well established as a central mediator of immune tolerance at the maternal-fetal interface through its interactions with inhibitory receptors expressed on natural killer cells, T cells, and macrophages, and comparable mechanisms have also been implicated in tumor immune evasion [15, 16]. In contrast, the immunological role of HLA-F is less clearly defined but appears to be both distinct from and complementary to that of HLA-G. Unlike HLA-G, which predominantly exerts tolerogenic effects via soluble and membrane-bound isoforms, HLA-F mainly exists as an open conformer and may function as a regulator or chaperone of other HLA class I molecules [31]. Emerging evidence suggests that HLA-F may modulate immune responses either independently or in coordination with HLA-G, thereby fine-tuning inhibitory signaling pathways at the maternal-fetal interface and within the tumor microenvironment. Whether HLA-F primarily amplifies HLA-G-mediated inhibitory signals or represents a distinct layer of immune regulation remains to be determined, and elucidating the functional interplay between these non-classical HLA molecules will be essential for a more comprehensive understanding of immune tolerance in both pregnancy and cancer.
With advancing gestational age, the expression of HLA-F at the maternal-fetal
interface gradually increases [19, 52]. HLA-F expression is lower in trophoblasts
compared to immune cells, and its protein levels decrease progressively during
differentiation from VCT to EVT, while HLA-G expression concurrently increases
[10]. Although HLA-F expression decreases during VCT-to-EVT differentiation,
functionally relevant levels of HLA-F remain detectable on EVTs. However, the
regulatory mechanisms underlying this dynamic shift in HLA expression during EVT
differentiation remain poorly studied. Previous and recent studies have
demonstrated that progesterone regulates HLA-G, HLA-E, and HLA-F expression in
EVTs [53, 54]. Other co-regulatory factors include IFN-
Transcription factors involved in EVT differentiation and HLA-G regulation, such as HIF-1A and GATA family proteins, could serve as intermediaries through which HLA-F influences EVT differentiation and HLA-G expression. It is possible that HLA-F promotes HLA-G expression by targeting transcription factors like HIF-1A and GATA2 via histone lactylation modifications.
Compared with HLA-G, HLA-F is more broadly expressed across multiple cell types at the maternal-fetal interface, a pattern similar to that observed in the TME. Consistent with this residual expression, HLA-F on EVTs is sufficient to engage inhibitory receptors on decidual immune cells and exert immunomodulatory effects. Single-cell transcriptomic analyses and immunohistochemistry have shown that HLA-F is widely and uniformly expressed in decidual immune cells, decidual stromal cells, VCTs, syncytiotrophoblasts, and EVTs [10, 20]. Overexpression of HLA-F in NK cells increases colony stimulating factor 1 (CSF1) secretion [10], and single-cell sequencing similarly reveals elevated CSF1 expression in decidual NK cells during early pregnancy [24]. CSF1 plays a key role in recruiting and inducing the differentiation of M2 macrophages [59, 60] and in promoting trophoblast invasion [61]. HLA-F on NK cells can also bind inhibitory receptors such as ILT2, ILT4, and VISTA on macrophages, thereby suppressing immune activation and maintaining immune tolerance at the maternal-fetal interface [18]. Beyond its potential role in inducing the expression of HLA-G, the “terminal effector” that sustains tolerance in decidual immune cells, HLA-F on EVTs and syncytiotrophoblast throughout pregnancy can bind inhibitory receptors such as KIR3DL1 and ILT2 on decidual or circulating NK cells, suppressing NK cytotoxicity. By engaging ILT2/4 and VISTA on macrophages and T cells, HLA-F further promotes early pregnancy immune tolerance and maintains immunological equilibrium at the interface.
A recent study demonstrated that HLA-F overexpression in Jar cells enhances glycolysis by upregulating PKM2 expression and inhibiting Pyruvate Kinase M2 Lysine 305 (PKM2 K305) lactylation, thereby increasing enzymatic activity and promoting cell proliferation [23]. This mirrors mechanisms observed in the TME, where tumor cells leverage a lactate-rich milieu to enhance their proliferative capacity [22] and reprogram immune cells through lactylation, suppressing NK- and TAM-mediated cytotoxicity to achieve immune escape [28, 34]. Decidual stromal cells also express HLA-F, and HLA-F expression in decidual tissue is associated with markers of normal decidual function and reproductive success. However, direct mechanistic evidence demonstrating a causal role of HLA-F in promoting decidualization is currently lacking. Several studies have shown that higher HLA-F expression in the decidua correlates with shorter time to pregnancy in assisted reproductive technology (ART) treatments [62, 63], potentially through recruitment of M2 macrophages [62], a mechanism reminiscent of immune-escape pathways in the TME. Although no direct mechanistic studies have yet confirmed that HLA-F enhances endometrial decidualization, the established role of HLA-F in promoting glycolysis in tumor cells suggests a plausible mechanism whereby elevated HLA-F enhances glycolysis to support decidualization. Multiple studies report that reduced glycolysis at the implantation site is associated with spontaneous miscarriage [64, 65], that hypoxia-induced decreases in embryonic lactylation impair mouse embryo development [66], and that diminished decidual glycolysis contributes to preeclampsia pathogenesis [67]. Collectively, these findings suggest that abnormal HLA-F expression may be linked to impaired fertility and pregnancy complications.
Several studies have reported that higher endometrial HLA-F expression during the mid-luteal phase is associated with shorter time-to-pregnancy in ART treatments, suggesting that elevated endometrial HLA-F levels enhance female fertility [62, 63]. Additionally, accumulating evidence suggests an association between altered HLA-F expression at the maternal-fetal interface and preeclampsia. Earlier studies using immunohistochemistry and western blotting observed significantly reduced HLA-F expression in EVTs from preeclampsia patients, though the underlying mechanisms were not thoroughly investigated [10, 20]. Subsequent research indicated that low HLA-F expression may inhibit trophoblast proliferation by decreasing glycolysis, thereby contributing to preeclampsia development [23]. Although one study validated the role of glycolysis in promoting proliferation by using a PKM2 activator in a MiniPDX model with transplanted shHLA-F-Jar cells, animal models with HLA-F knockout remain lacking to confirm HLA-F’s definitive role in fertility.
Aside from individual variations in transcription factors such as NF-
In support of these findings, Langkilde CH et al. [63] performed Sanger sequencing in a genetic analysis of 102 infertile women undergoing ART and 78 controls. They identified significant associations between longer time-to-pregnancy, measured as the number of ART cycles required, and HLA-F genotypes linked to lower mid-luteal endometrial HLA-F mRNA expression, specifically the rs1362126 AA, rs2523405 GG, and rs2523393 GG genotypes [63]. Follow-up validation studies involving a prospective cohort of 85 recurrent implantation failure (RIF) patients and 36 controls, alongside primary endometrial culture experiments, confirmed that these three HLA-F gene polymorphisms influenced endometrial HLA-F protein expression. Furthermore, they reported positive correlations between endometrial HLA-F expression, anti-inflammatory CD163+ M2 macrophage infiltration, and CD56+ uterine NK cell abundance [62]. Recently, a genotyping study by qPCR involving 164 preeclampsia patients, 19 fetal growth restriction (FGR) patients, and 192 healthy controls identified that the frequency of the rs2523393 A allele, an HLA-F eQTL SNP, was lower in preeclampsia patients (55.8%) compared to healthy controls (58.9%), although this difference did not reach statistical significance. Nevertheless, a trend toward the absence of certain HLA-F–HLA-G extended diplotypes in preeclampsia was noted [71].
However, whether existing studies on HLA-F polymorphisms and their associations with fertility or preeclampsia have included sufficiently large sample sizes to ensure robust conclusions remains questionable. Research on HLA-G polymorphisms and preeclampsia is the most extensive, yet findings remain inconsistent [49, 71, 72]. Studies examining HLA-F polymorphisms and preeclampsia are even more limited, with only a single study involving 164 cases reported to date [71]. For instance, assuming an HLA-F*01:01:02 allele frequency of approximately 16.62% [73, 74, 75], and a preeclampsia incidence of 3%, one would expect ~150 disease cases to detect an odds ratio (OR) of 0.4 (a 60% risk reduction). However, given that preeclampsia is widely regarded as a multifactorial, multigenic disorder, it is unlikely that a single gene contributes such a large effect. If the true effect size corresponds to a 20% reduction in risk (OR = 0.8), approximately 2400 cases would be required. Therefore, inconsistencies in studies examining polymorphisms in non-classical HLA-Ib genes and their relationships with fertility or preeclampsia likely stem from underpowered sample sizes. Moreover, synergistic effects may exist among HLA-Ib molecules; disease associations may depend on haplotype combinations, as reported for coronary artery disease, which is linked to HLA-F/G/E polymorphism combinations [76], and preeclampsia, which may be associated with specific HLA-F–HLA-G haplotypes [71]. Given the small sample sizes of existing studies, large multicenter cohorts are necessary to clarify the relationships between HLA-F polymorphisms or HLA-Ib haplotypes and disease susceptibility.
Although several studies have demonstrated that elevated HLA-F protein or LncRNA HLA-F-AS1 expression in tumors correlates with poor prognosis [7, 25], research investigating associations between HLA-F genetic polymorphisms and cancer incidence remains scarce. Such studies could have significant implications for understanding tumor genetic susceptibility. In viral infections, the HLA-F*distal-D haplotype, harboring the 1610 C allele and associated with lower HLA-F mRNA expression compared with the HLA-F*distal-C haplotype, showed an increased frequency among Zika virus (ZIKV)-, Dengue virus (DENV)-, or Chikungunya virus (CHIKV)-infected patients diagnosed with peripheral spectrum disorders (which generally have better clinical outcomes than encephalitic spectrum disorders) [14]. Another study found that children carrying the G allele at HLA-F (–1499, rs183540921) exhibited an increased asymptomatic/symptomatic ratio of Plasmodium falciparum malaria [15]. In addition, the HLA-F*01:03 allele, associated with lower mRNA expression than HLA-F*01:01, has been linked to reduced hepatitis B virus (HBV) DNA levels [77]. These findings suggest that, unlike in reproduction, lower HLA-F expression may confer better outcomes following viral infection, whereas higher expression may worsen infection-induced complications.
In autoimmune diseases, however, higher HLA-F–expressing genotypes appear to have protective effects. In ankylosing spondylitis (AS), the allele HLA-F*01:01:02 (associated with higher mRNA expression) (p = 0.0049; pc = 0.0098; OR = 0.60) and its corresponding SNP rs2075682 (p = 0.0004; pc = 0.0008; OR = 0.53) were suggested to be protective [78]. Conversely, the rs2523393 G allele (associated with lower mRNA expression) has been linked to malabsorption, celiac disease, multiple sclerosis, and hyperthyroidism [53]. In Systemic Lupus Erythematosus (SLE), anti–HLA-F IgG may participate in clearing HLA-F shed from lymphocytes and inflamed tissues, potentially mitigating disease severity and serving as a beneficial immune biomarker [79]. Ravindranath et al. [80] further showed that antibodies targeting HLA-F heavy chains were the predominant anti-HLA-Ib heavy-chain IgG antibodies in patients with rheumatoid arthritis (RA) and systemic lupus erythematosus (SLE). However, these studies did not evaluate whether anti-HLA-F IgG levels correlate with HLA-F genetic polymorphisms.
Collectively, current evidence indicates that HLA-F plays opposing roles in reproduction and disease. Genotypes associated with higher HLA-F expression appear beneficial for fertility and immune regulation but may worsen outcomes in cancer and viral infections. For example, the rs2523393 A allele, thought to have arisen in recent human evolution [53], is associated with higher HLA-F expression, enhanced fecundability, reduced preeclampsia risk, and lower incidence of autoimmune diseases such as multiple sclerosis and hyperthyroidism. However, this same allele may predispose carriers to increased tumor metastasis and poorer cancer prognosis. Given that HLA-F polymorphisms have remained highly conserved throughout human evolution, it is plausible that this gene embodies an evolutionary tradeoff, a reproduction–health balance maintained by antagonistic pleiotropy, wherein variants that promote reproductive success simultaneously elevate long-term health risks such as cancer progression.
Several limitations of this review should be acknowledged. The current conclusions are largely derived from observational and associative studies, and direct functional evidence defining the causal role of HLA-F remains limited. In addition, heterogeneity in study design, population characteristics, and analytical approaches across the literature may contribute to variability in reported findings. Furthermore, although parallels between the immunological roles of HLA-F in pregnancy and cancer are increasingly recognized, mechanistic links between these two contexts remain incompletely defined. Addressing these limitations through integrated experimental and clinical studies will be essential to further clarify the immunoregulatory functions of HLA-F.
Numerous studies have demonstrated that elevated expression of HLA-F protein or lncRNA HLA-F-AS1 correlates with poor prognosis across various malignancies. However, mechanistic investigations into the causes of HLA-F upregulation and studies examining associations between HLA-F polymorphisms and cancer incidence are currently lacking. Investigating genetic polymorphisms in HLA-F may help identify novel tumor susceptibility markers, while further research into the role of HLA-F in immune escape within the TME could reveal new therapeutic targets for cancer immunotherapy.
Based on existing evidence, elevated HLA-F expression at the maternal-fetal interface appears to be beneficial for reproduction, as reflected by shorter time-to-pregnancy in assisted reproductive treatments and its downregulation in pregnancy complications such as preeclampsia [10]. Conducting large-scale studies to investigate relationships between pregnancy complications and HLA-F polymorphisms or HLA-Ib haplotype frequencies could uncover novel genetic markers associated with fertility, preeclampsia, and other pregnancy complications. Moreover, elucidating the mechanisms underlying HLA-F-mediated immune tolerance at the maternal–fetal interface may offer innovative strategies and therapeutic interventions to enhance human fertility and reduce pregnancy-related complications.
LJ drafted the initial version and created the sketches. LJ, FL and JW designed of the work and revised the draft. All authors contributed to editorial changes in the manuscript. All authors have read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
No applicable
This research received no external funding.
The authors declare no conflicts of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.