






 , Ying Lyu 2, E Fen Cheng 2
, Ying Lyu 2, E Fen Cheng 21 Central Laboratory and Precision Medicine Center, Affiliated Jinhua Hospital, Zhejiang University School of Medicine, 321000 Jinhua, Zhejiang, China
2 Departments of Obstetrics and Gynecology, Affiliated Jinhua Hospital, Zhejiang University School of Medicine, 321000 Jinhua, Zhejiang, China
Abstract
Although multiple factors contribute to recurrent spontaneous abortion (RSA), recent studies have highlighted a role for regulatory T cells (Tregs) in maintaining immune tolerance during pregnancy. Understanding the intricate relationship between RSA and Tregs offers promising avenues for diagnosis and therapeutic strategies in reproductive medicine.
We successfully established a mouse model of RSA, and all animals were euthanized on gestational day 13. Uterine horns were examined upon dissection to determine the number of viable fetuses and resorption sites, after which the post-implantation loss rate (abortion rate) was calculated. Placental morphology was evaluated via hematoxylin and eosin staining, whereas gene expression was analysed by immunohistochemistry. Treg abundance was determined by flow cytometry. Gene expression and cytokine levels in EL4 cells and patient samples were evaluated by quantitative real-time polymerase chain reaction (qPCR) and Western blotting.
The abortion rate of RSA mice was 31.64%. In addition, Treg numbers were reduced in both RSA mice and patients, while the expression of Toll-like receptor 4 (TLR4) and p65 was upregulated, further suppressing forkhead box protein P3 (FOXP3) expression. Blocking p65 expression with small interfering RNA (siRNA) targeting p65 prevented the lipopolysaccharide (LPS)-induced downregulation of FOXP3 in EL4 cells. The TLR4 inhibitor IAXO102 was ineffective at increasing FOXP3 expression in EL4 cells following p65 overexpression p65.
The results of this study suggest that activation of the TLR4/p65 signalling in RSA inhibits FOXP3 expression and contributes to abortion. Importantly, the findings indicate that Treg suppression in RSA is p65-dependent.
Graphical Abstract
Keywords
- recurrent spontaneous abortion
- regulatory T cells
- forkhead box P3
- toll-like receptor 4
- nuclear factor kappa-B
Recurrent spontaneous abortion (RSA), or recurrent miscarriage, is defined as three or more consecutive pregnancy losses prior to 28 weeks’ gestation [1]. This condition presents substantial physical and emotional challenges, necessitating a multidisciplinary approach to care [2]. Advances in our understanding of the mechanisms involved, particularly those relating to maternal immune dysregulation and cytokine imbalances, may reveal novel therapeutic targets and improve outcomes [3]. Continued research is essential to develop effective interventions and enhance reproductive health.
Regulatory T cells (Tregs) are essential for maintaining peripheral tolerance, preventing autoimmunity, and establishing fetal tolerance during pregnancy [4, 5]. A reduction in Treg number and function mediates fetal rejection, a phenomenon that is reversible in animal models via Treg transplantation [6]. Forkhead box protein P3 (FOXP3), a key transcriptional regulator of Treg development, has also been associated with reproductive failure when underexpressed in endometrial tissue [7]. This study found that peripheral blood Treg levels were significantly decreased in patients with RSA.
Toll-like receptor 4 (TLR4) recognises pathogen-associated molecular patterns,
such as lipopolysaccharide (LPS), initiating signalling that causes nuclear
translocation of the nuclear factor kappa-light-chain-enhancer of activated B
cells (NF-
To explore the role of immune dysregulation in RSA, we used a mouse model and human specimens. We discovered that the TLR4/p65 pathway is hyperactive in RSA and directly inhibits FOXP3, which is a key regulator of protective Tregs. By using a pathway agonist (LPS) and antagonist (IAXO102), as well as genetically knocking down and overexpressing p65, we demonstrated, at a mechanistic level, that targeting this pathway modulates FOXP3 expression. These findings reveal a novel therapeutic strategy for preventing miscarriage.
This study recruited 20 patients with RSA and 20 healthy control participants. Inclusion criteria for the RSA group: Females aged 20–45 years who meet the criteria for an RSA diagnosis, with a current intrauterine pregnancy confirmed via ultrasound. Both partners must provide informed consent and sign the consent form. Exclusion criteria included known causes of miscarriage, anatomical abnormalities of the reproductive tract, uncontrolled endocrine or metabolic disorders, autoimmune diseases, prothrombotic states, recent acute infections and other major systemic diseases. The control group comprised healthy women in early pregnancy who had voluntarily terminated their pregnancy for non-medical reasons (the normal early pregnancy abortion group). The gestational age of this group was matched with that of the RSA group, and all the RSA group’s exclusion criteria were applied. All participants provided informed consent and signed informed consent forms.
This study involved collecting products of conception tissue and peripheral blood.
On the day of the ultrasound-guided abortion or surgical procedure, the tissue was obtained using sterile instruments and immediately divided into two portions. One portion was placed in 4% paraformaldehyde for fixation, followed by paraffin embedding, histological sectioning and immunohistochemical analysis. The other portion was placed in a sterile, enzyme-free cryopreservation tube, rapidly frozen in liquid nitrogen and transferred to –80 °C storage for subsequent molecular biology experiments, such as protein/RNA extraction, Western blot and quantitative real-time polymerase chain reaction (qPCR). Peripheral blood: Collect 5–10 mL of fasting venous blood from patients in the morning using EDTA anticoagulant tubes (Bacet, Wenzhou, Zhejiang, China). Centrifuge at 3000 rpm for 15 minutes at 4 °C. Aspirate the plasma from the supernatant, divide it into smaller portions, and store it at –80 °C. All sample tubes were labelled with unique, traceable codes (concealing patient names) and a sample information database was created. This database recorded the following information for each sample: ID, patient group, age, parity, number of miscarriages, gestational age, collection date, processing method and storage location. Biological samples were stored in –80 °C freezers or liquid nitrogen tanks, and storage equipment operation was regularly checked.
A total of forty-five 6-week-old mice, comprising thirty CBA/J, five DBA/2, ten
BALB/c, were purchased from GemPharmatech™
(Nanjing, Jiangsu, China) and housed in stainless steel cages with ad libitum access to
food and water. They were kept under controlled conditions of temperature (22
The EL4 cells were obtained from the Cell Bank of the Chinese Academy of Sciences (TCM41, Shanghai, China). The cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) (Gibco, Waltham, MA, USA) at 37 °C in a humidified atmosphere containing 5% CO2. The medium was refreshed every three days, and the cells were subcultured when they reached 80% confluence. Mycoplasma testing was negative. The EL4 cells have been authenticated by STR profiling.
The uterine tissue segments were fixed in 4% paraformaldehyde, dehydrated using a graded ethanol series and embedded in paraffin wax Thin sections were stained with haematoxylin and eosin (H&E) and examined using the Zeiss light microscope (Oberkochen, Germany). For immunohistochemistry, the placental sections were deparaffinised and treated with 3% hydrogen peroxide to block endogenous peroxidase. They were then incubated with goat serum to minimise non-specific binding. An overnight incubation of the slides was performed at 4 °C with anti-TLR4 antibody (1:500, 19811-1-AP; proteintech, Wuhan, Hubei, China), anti-p65 antibody (1:200, 3033; Cell Signaling Technology, Danvers, MA, USA), and anti-FOXP3 antibody (1:500, 22228-1-AP; proteintech, Wuhan, Hubei, China). After three washes in 0.01 M PBS, the samples were incubated with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (1:5000, TA373083L; OriGene, Beijing, China) for 30 min at 37 °C. Immunostaining was visualised using 3,3′-diaminobenzidine (DAB) and the sections were counterstained with Mayer’s haematoxylin (ZSBG-BIO, Beijing, China). The sections were then dehydrated, cleared and mounted. Immunohistochemical staining for TLR4, p65 and FOXP3 was quantified using a standardised H-scoring system [19].
The frequency of Tregs was determined using flow cytometry to analyse single-cell suspensions from mouse spleens and human peripheral blood. Spleens were harvested from each group and processed into single-cell suspensions using a previously established protocol [20]. Human peripheral blood mononuclear cells were isolated using Ficoll density gradient centrifugation. To quantify the CD4+CD25+FOXP3+ T cell population, mouse and human Tregs staining kits (88-8111-40 and 88-8999-40, respectively, eBioscience, San Diego, CA, USA) were employed in accordance with the manufacturers’ instructions. Data are presented as the percentage of Tregs within the total cluster of differentiation 4 (CD4)+ T cell population [21].
Total protein was extracted from the abortion products of patients with RSA, and the protein concentration was determined by the bicinchoninic acid (BCA) method. Immunoblotting was performed using standard methods. TLR4 antibody (1:500, 19811-1-AP; proteintech, Wuhan, Hubei, China), TLR4 (1:2000, ab13556; Abcam, Cambridge, UK), p65 (1:1000, 3033; Cell Signaling Technology, Danvers, MA, USA), and FOXP3 (1:2000, ab215206; Abcam, Cambridge, UK) were used to detect the proteins. GAPDH (1:50,000, 60004-1-Ig, proteintech, Wuhan, Hubei, China) was used as an internal control to normalise protein loading. Anti-rabbit (1:5000) secondary antibodies were conjugated with HRP (P6782; Sigma-Aldrich, Shanghai, China). HRP-labeled Goat Anti-Mouse IgG (H + L) (1:2000, A0216, Beyotime, Shanghai, China) were used. Following separation, the membranes were trimmed according to the migration of a colorimetric protein marker. Immunoreactive bands were detected using an enhanced chemiluminescence system (Merck, Darmstadt, Germany) and quantified using densitometry with ImageJ software (Version 1.54; National Institutes of Health, Bethesda, MD, USA).
Total RNA was extracted from the abortion products of patients with RSA using
Trizol reagent (15596026; Invitrogen, Carlsbad, CA, USA) for qPCR analysis. The
expression of three target genes (TLR4, p65, FOXP3) and the housekeeping gene
(
Small interfering RNAs (siRNAs) were synthesised by Genepharma (Shanghai, China). A universal control siRNA was used as a non-specific control. EL4 cells were transfected with the siRNA duplex using Lipofectamine 2000 (Invitrogen) according to the manufacturer’s instructions. The p65 genes were amplified and cloned into a pcDNA3.1 vector (Novagen, Madison, WI, USA). Transfection of EL4 cells was achieved using the Amaxa Nucleofector system (Thermo Fisher Scientific, Waltham, MA, USA).
Statistical analyses were performed using GraphPad Prism 8 software (GraphPad Software, Inc., San Diego,
CA, USA). Student’s t-test was used for intergroup comparisons,
and one-way ANOVA was used for multiple comparisons. Tukey’s post-hoc test was
then used to identify specific differences between groups. A p-value of
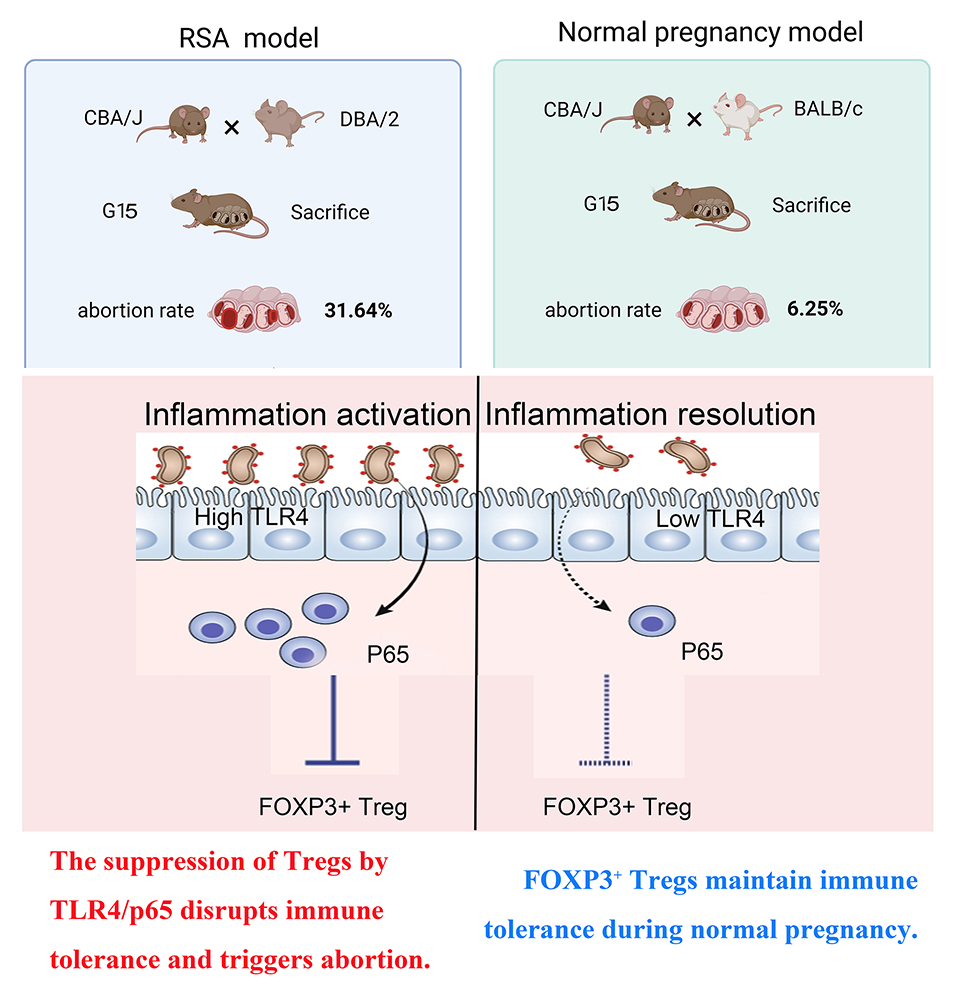
To evaluate the divergence in gestational outcomes in RSA mice, both normal control and RSA pregnancy models were established, as previously described [22]. In the normal pregnancy group, mice received intraperitoneal injections of either PBS or RU486 during early gestation (G5). All mice were euthanised at G13. In the normal pregnancy model, administration of RU486 induced an abortion rate of 97.53%, which was markedly higher than the 6.25% observed in the PBS-treated control group, confirming the efficacy of RU486 in establishing an early pregnancy loss model. However, the miscarriage rate in the RSA pregnant mouse model was 31.64% (Table 1). The RSA group experienced fetal and placental necrosis and haemorrhage, whereas control mice had almost no obvious fetal abnormalities (Fig. 1A). H&E staining was performed on uterine sections from the control, RU486-treated, and RSA model groups (Fig. 1B). The maternal–fetal interface in the RSA mouse model displayed pronounced haemorrhagic changes and inflammatory characteristics. This result confirmed that the RSA model was successfully constructed.
| Group | Number of mice | Surviving fetuses | Resorbed fetuses | Resorption rate (%) |
| Control | 10 | 75 | 5 | 6.25 |
| RU486 | 10 | 2 | 79 | 97.53*** |
| RSA | 10 | 54 | 25 | 31.64* |
The abortion rate was determined by dividing the number of abortion sites by the
total number of implantation sites. Group comparisons for abortion rates were
performed using Student’s t-test. Statistical differences between groups
are shown as follows: Compared with the control group, *p
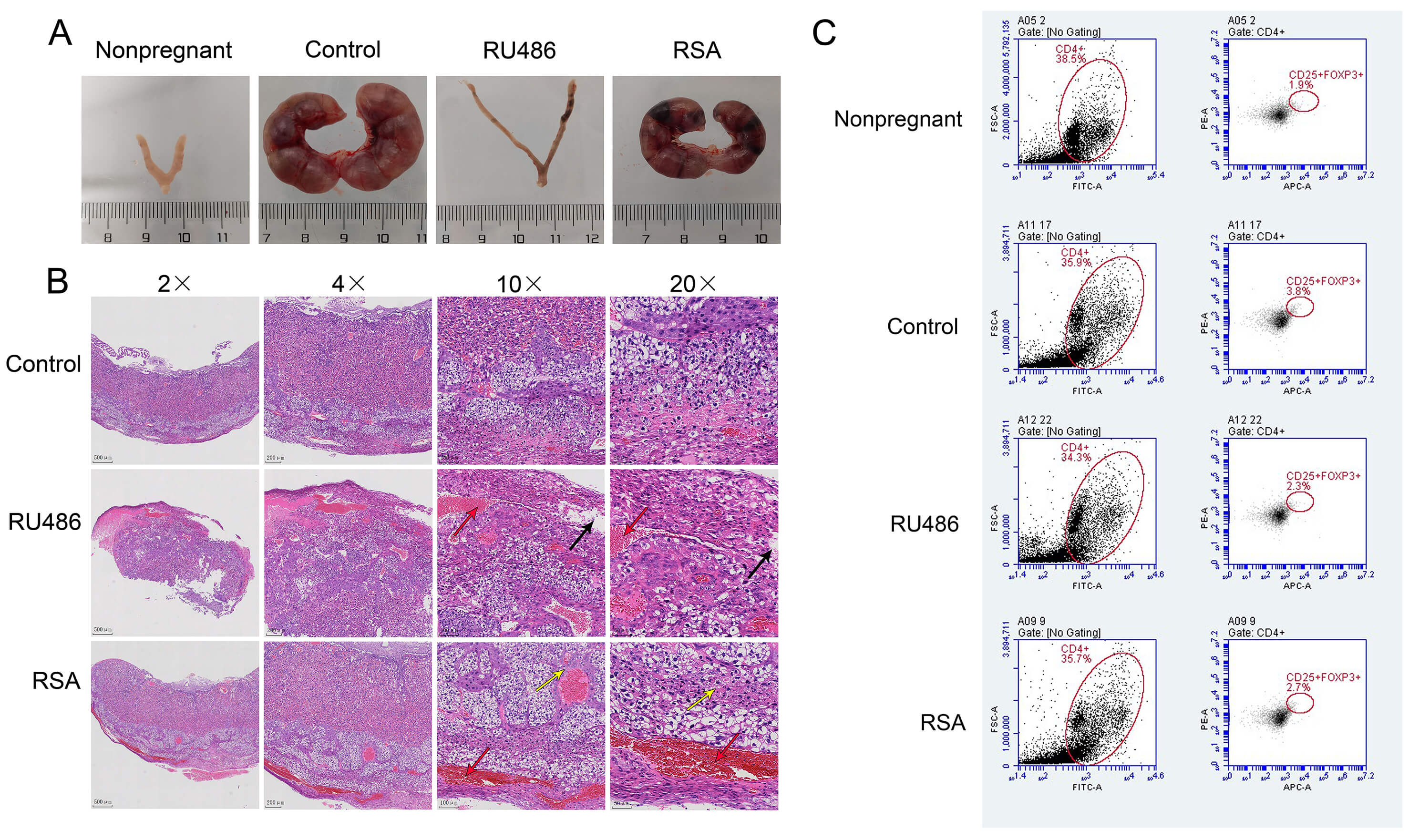 Fig. 1.
Fig. 1.
Reduced Treg cell expression in RSA mice. (A)
Representative uterine specimens were collected from both normal and RSA-affected
pregnancies at G13. (B) Histopathological evaluation using H&E staining was
performed on maternal–fetal interface sections from the corresponding
experimental groups. The placental structure was clearly defined in the RSA
group, but disorganised in the RU486 group. Black arrows mark loose oedema,
yellow arrows indicate necrotic cells, red arrows indicate bleeding. 2
Given the established role of CD4+CD25+FOXP3+ T cells in maternal–fetal immune tolerance, their prevalence among splenocytes was quantified using flow cytometry. The proportion of these Tregs was significantly reduced in both the RSA and RU486 groups. However, it was still higher than that of the non-pregnant group (Fig. 1C). The statistical graph for Fig. 1C is shown in Supplementary Fig. 2. These findings suggest that impaired expansion or maintenance of CD4+CD25+FOXP3+ T cells is associated with pregnancy loss in the RSA model.
FOXP3, a pivotal transcription factor governing the development and function of
Tregs, plays a critical role in diverse immunological processes including immune
homeostasis, autoimmune pathogenesis, infectious responses, and tumour immune
evasion [23]. The expression of TLR4, p65, and FOXP3 in placenta tissue was
assessed using Western blotting and immunohistochemistry. As shown in Fig. 2
(p
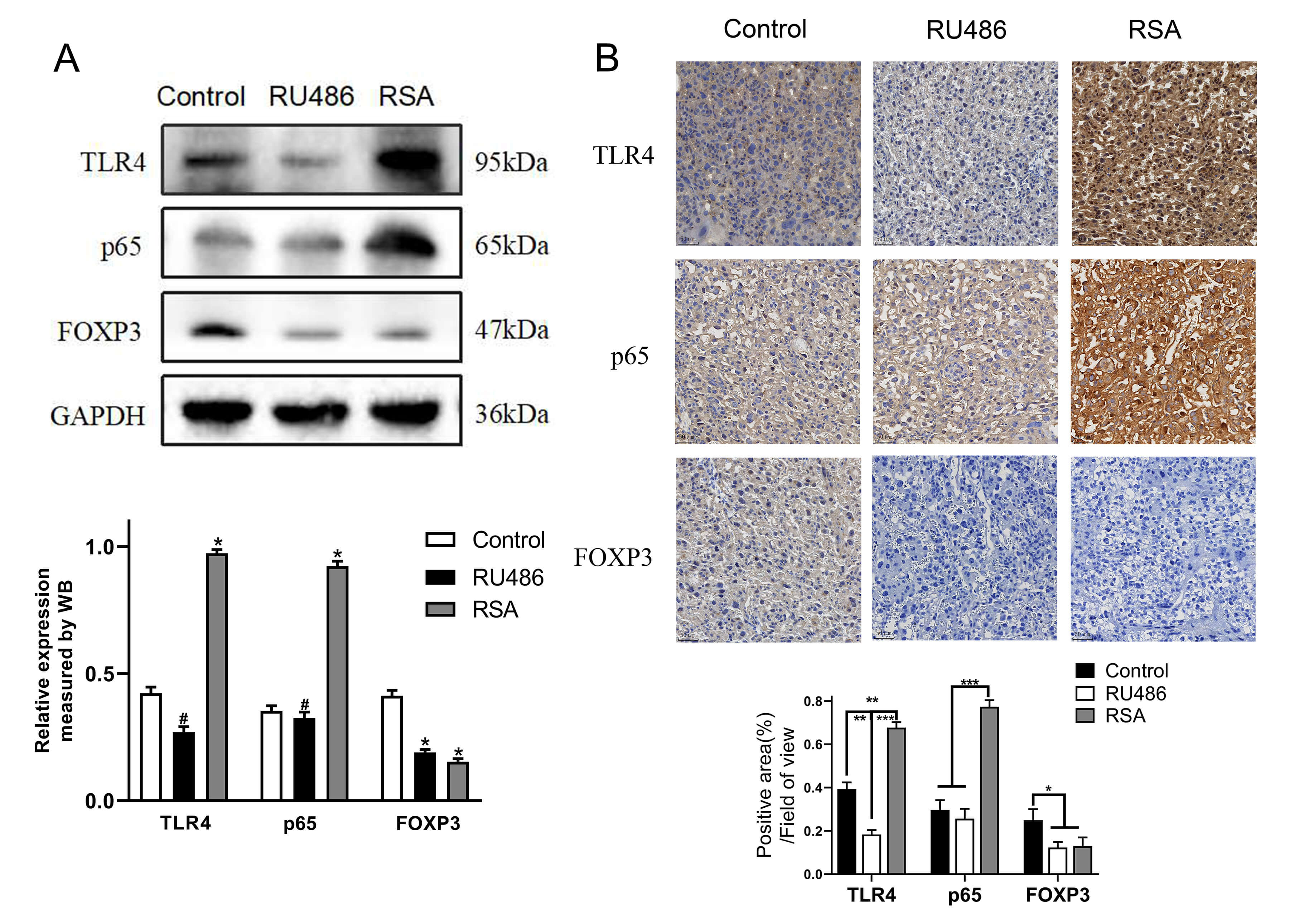 Fig. 2.
Fig. 2.
Regulation of the TLR4/p65/FOXP3 signalling pathway in RSA
mice. (A) The protein expression levels of TLR4, p65, and FOXP3 in placental
tissue were assessed by Western blotting. (B) TLR4, p65, and FOXP3 was detected
by immunohistochemistry. Scale bar: 50 µm. *: Compared with the
control group, p
Activation of the TLR4 signalling pathway significantly contributes to the
downregulation of FOXP3 expression [24]. To verify the link between RSA and Treg
reduction, Treg levels in the peripheral blood of patients with RSA were assessed
using flow cytometry. Compared with the control group, the number of Tregs in
patients with RSA was reduced (Fig. 3A and Supplementary Fig. 3,
p
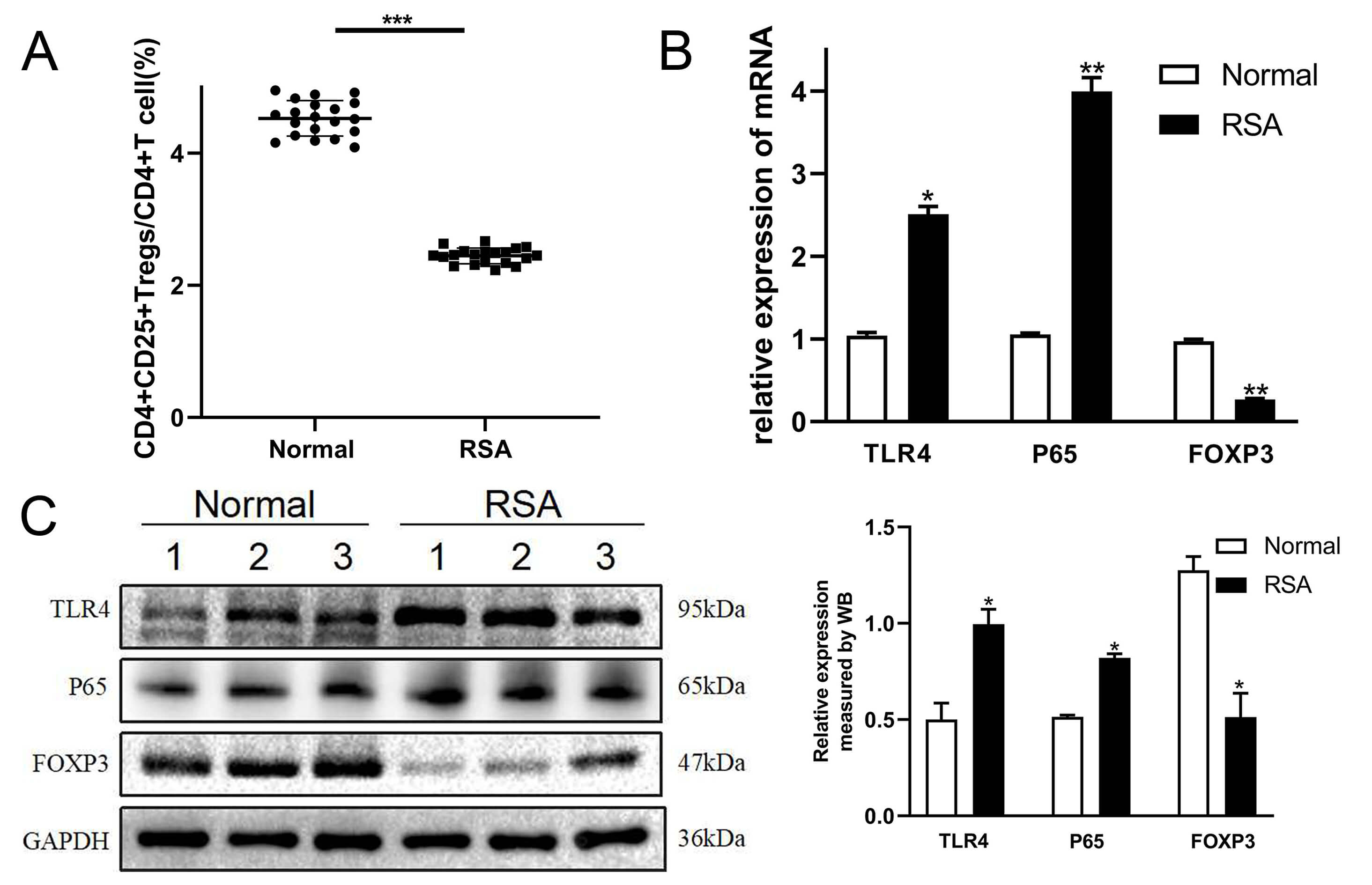 Fig. 3.
Fig. 3.
Expression levels of TLR4, p65, and FOXP3 in patients with RSA.
(A) Percentage of CD4+CD25+FOXP3+ T cells in peripheral blood was assessed using
flow cytometry. (B) qPCR was performed to measure TLR4, p65, and FOXP3 transcript
levels in decidual tissue. (C) Protein expression of TLR4, p65, and FOXP3 were
detected by Western blotting. *: compared with the normal decidua, p
To investigate the underlying molecular mechanisms, we employed the EL4 cell
line, a model characterised by strong inducible FOXP3 expression upon
stimulation, while retaining numerous intrinsic T-cell properties [25]. To assess
the involvement of p65 in RSA-induced FOXP3 downregulation, we initially
stimulated the TLR4/p65 pathway using the agonist LPS, and subsequently inhibited
p65 expression via siRNA targeting p65 (si-p65), with a non-targeting control
siRNA (si-NC) serving as the negative control. Moreover, the TLR4 inhibitor
IAXO102 was chosen to inhibit the TLR4/p65 signalling pathway, followed by
overexpression of p65 with pcDNA3.1-p65, the corresponding empty vector control
(pcDNA-NC) was used as the negative control. Compared to the
si-NC group, transfection with si-p65 significantly reduced p65 protein
expression (Fig. 4A, p
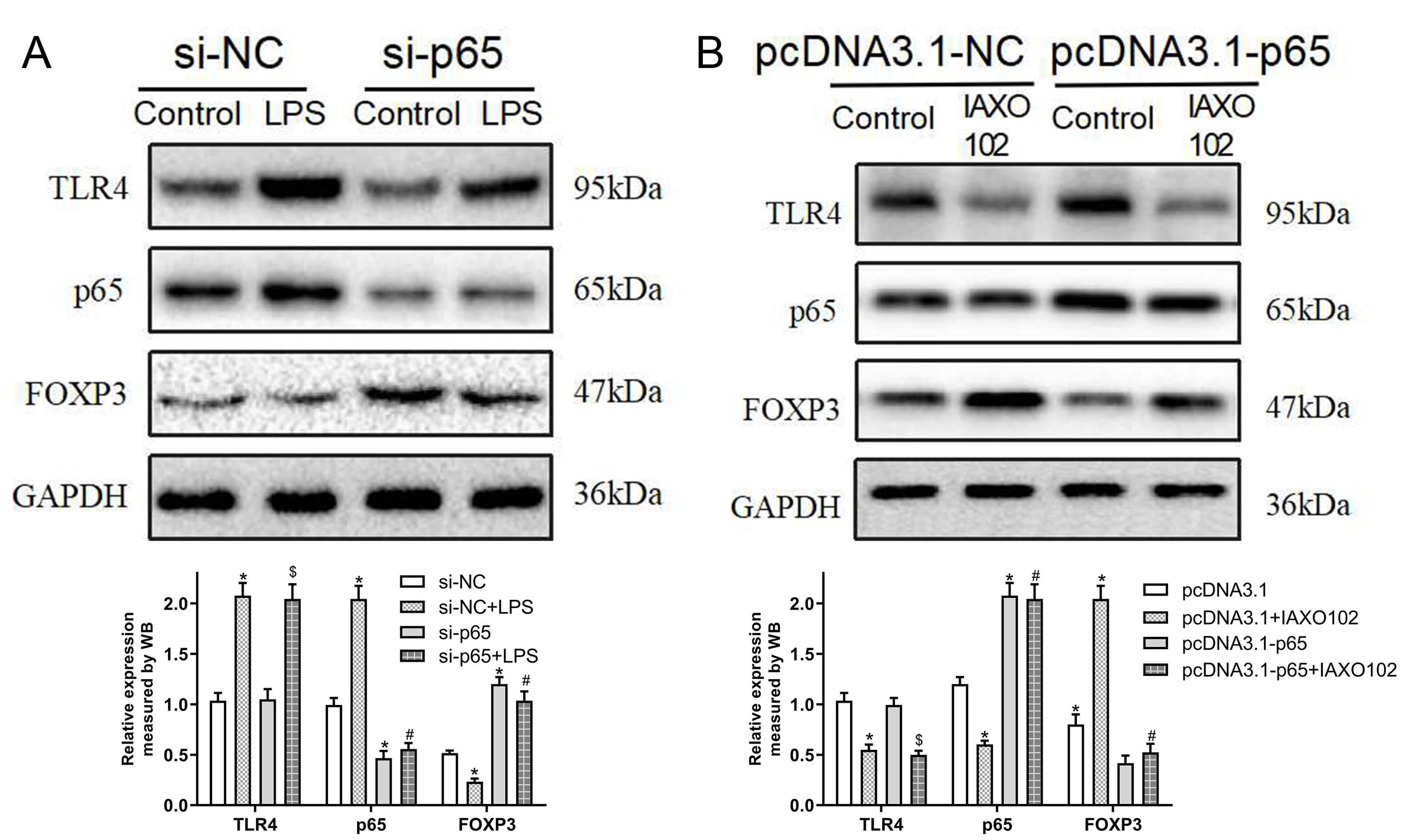 Fig. 4.
Fig. 4.
Downregulation of FOXP3 by TLR4 signalling is p65-dependent.
(A) EL4 cells were transfected with p65-target siRNA or control siRNA with or
without LPS (TLR4 agonist). Western blotting showed TLR4, p65, and FOXP3 protein
expression. *p
RSA is a multifactorial disorder that involves genetic, anatomical, hormonal, immunological and environmental factors [26]. Further research into its aetiology is essential for improving diagnosis, risk assessment, and clinical management [27]. Furthermore, the substantial medical and psychological burden imposed by RSA highlights the need for further investigation to alleviate its impact on patients and healthcare systems [28].
In the case of RSA, disturbances of the immune tolerance mechanism may lead to miscarriage [29]. Recent research identified Tregs as a subpopulation with immunosuppressive functions [30]. FOXP3 serves as a lineage-defining transcription factor that governs the development and suppressive function of Tregs. Treg-mediated immunoregulation is critically involved in maintaining self-tolerance and preventing autoimmune pathologies, as well as establishing maternal–infant immune tolerance during pregnancy [31]. During normal pregnancy, the frequency of Tregs in peripheral blood rises during early pregnancy. These cells subsequently migrate to the maternal–infant interface, where they respond to various chemokines produced by human chorionic gonadotropin and trophoblast cells [32]. Consistent with previous findings in both animal models and humans, our results reconfirm that Treg depletion is associated with maternal immune dysregulation in conditions such as RSA [33]. We observed a reduction in Tregs in the RSA mouse model, aligning with earlier reports of suppressed Treg levels in the peripheral blood of patients with RSA.
As a specific TLR4 agonist, LPS activates TLR4 and its downstream NF-
To further investigate the regulation of FOXP3 by p65, we used p65-specific
siRNA and overexpression plasmids. We found that p65 knockdown prevented
LPS-induced downregulation of FOXP3, and p65 overexpression inhibited
IAXO102-induced FOXP3 upregulation. These results suggest that p65 directly
regulates FOXP3 expression in this model, providing mechanistic support for the
notion that TLR4/NF-
A key limitation of this study is the use of the EL4 cell line, which may not
fully replicate the complexity of the maternal-fetal immune microenvironment
in vivo. Furthermore, extrapolating these findings to human clinical
contexts requires validation through in vivo models and human tissue
studies. Future work should also explore the interactions between trophoblasts
and immune cells within pathological microenvironments to better understand the
potential for translating TLR4/NF-
In summary, this study reconfirms the association between Treg deficiency and
recurrent miscarriage, suggesting that the TLR4/NF-
All data points generated or analyzed during this study are included in this article and there are no further underlying data necessary to reproduce the results.
CH designed the research study and performed the research. EFC provides assistance and guidance for clinical specimen collection, making substantial contributions to data acquisition and analysis. YL analyzed the data. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
This study was reviewed and approved by the Medical Ethics Committee of Jinhua Municipal Central Hospital (Approval No. 2023-149; Jinhua, Zhejiang Province, China). The informed consent was obtained. The study was conducted in accordance with the principles of the Declaration of Helsinki. All animal experiments were conducted in accordance with institutional guidelines and the principles of the 3Rs (Replacement, Reduction, and Refinement). All animal experiments were approved by the Laboratory Animal Welfare and Ethics Committee of Jinhua Municipal Central Hospital (Approval No. AL-JHYY202327), and all animal experiments were performed following approved animal care and use protocols.
We would like to express our gratitude to all those who helped us during the writing of this manuscript. Thanks to all the peer reviewers for their opinions and suggestions.
This work was supported by Jinhua Science and Technology Bureau (2019-3-006), Medical Science and Technology Project of Zhejiang Province (2022KY1326) and Zhejiang Traditional Chinese Medicine Administration (2024ZL1182).
The authors declare no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/CEOG45730.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.