



 , Pingping Jiang 1,*
, Pingping Jiang 1,*
1 Department of Traditional Chinese Medicine, The First Affiliated Hospital of Guangdong Pharmaceutical University, 510060 Guangzhou, Guangdong, China
2 State Key Laboratory of Oncology in South China, Sun Yat-Sen University Cancer Center, 510060 Guangzhou, Guangdong, China
†These authors contributed equally.
Abstract
Yam Polysaccharide (YP) is a major bioactive component extracted from the common yam rhizome and has been shown to have an antidiabetic effect. Previous research has demonstrated the association between gut microbiota (GM) and gestational diabetes mellitus (GDM); however, whether GM is essential for mediating the antidiabetic effects of YP in GDM remains unclear.
A high-fat and fructose diet (HFD) was administered to mice before and after pregnancy to induce GDM. An oral glucose tolerance test (OGTT) was performed, and the homeostatic model assessment of insulin resistance (HOMA-IR) was calculated to evaluate glucose metabolism. The GM composition was analyzed using 16S rRNA sequencing.
During pregnancy, mice fed an HFD exhibited significant weight gain accompanied by impaired glucose tolerance. These metabolic disturbances were alleviated by YP treatment. Furthermore, consuming an HFD induced marked alterations in GM diversity compared to the control group, characterized by increased abundance of Alloprevotella and decreased abundance of the Lachnospiraceae NK4A136 group. Importantly, YP administration reversed these HFD-induced microbial changes.
These findings suggest that modulation of the GM is one mechanism underlying the antidiabetic effects of YP in GDM.
Keywords
- gestational diabetes mellitus
- gut microbiota
- 16S rRNA sequencing
- Yam Polysaccharide
Gestational diabetes mellitus (GDM) is defined as any degree of glucose intolerance that is recognized during pregnancy [1]. A global epidemiology study reports a prevalence of GDM at 17.8% [2]. Additionally, a meta-analysis involving 79,064 Chinese participants found an incidence rate of 14.8% [3]. GDM increases the risks of complications such as eclampsia, macrosomia, induced labor, preterm delivery, and cesarean section [4]. Moreover, pregnant women with GDM and their offspring are at a higher risk of developing type 2 diabetes mellitus (T2DM) [5, 6] and are more susceptible to cardiovascular diseases [7, 8] in the long term. Thus, the prevention and management of GDM has become one of the most important public health issues that needs to be addressed.
The mechanisms underlying the onset and progression of GDM have not been fully understood. Increasing evidence suggests that an imbalance of the gut microbiota (GM) may play a significant role in the development of metabolic disorders, obesity, and diabetes. GM is essential for host physiology, as it regulates nutrient metabolism, maintains the integrity of the intestinal barrier, modulates immune responses, and produces bioactive metabolites such as short-chain fatty acids (SCFAs). Through these functions, GM contributes to energy homeostasis, systemic immunity, and overall metabolic health. It has been observed that the composition of GM changes during pregnancy. A prospective cohort of 91 Finnish women found that GM obtained in the third trimester showed increased diversity but reduced richness. There was also a notable increase in Proteobacteria in pregnant women compared to non-pregnant controls [9]. Crusell et al. [10] demonstrated that GM composition in women with GDM, during and after pregnancy, resembled the altered microbiota seen in non-pregnant individuals with T2DM. Genera such as Desulfovibrio and Collinsella are enriched in patients with GDM and T2DM [10], with Collinsella specifically linked to higher fasting insulin levels and homeostatic model assessment for insulin resistance (HOMA-IR) [11]. Furthermore, bacteria such as Bacteroides dorei [12], Blautia [13], Klebsiella variicola [14], and Ruminococcaceae [15] have been identified as risk factors for GDM, while Bifidobacterium [14], Akkermansia [16], and Faecalibacterium [17] are considered protective. These bacteria play active roles in glucose and lipid metabolism and are closely associated with insulin resistance, potentially explaining how changes contribute to the development of GDM.
Chinese yam (Dioscorea opposita) has been used for centuries in traditional Chinese medicine and as a functional health food. Yam Polysaccharide (YP), the primary bioactive component found in the Chinese yam, is a high-molecular-weight polymer of monosaccharides (such as glucose, galactose, and mannose linked together in a complex structure. YP can be industrially extracted and purified, exhibiting multiple biological activities, including antioxidant, anti-inflammatory, and metabolic regulatory effects. These properties make YP a crucial compound for examining the therapeutic potential of Chinese yams [18]. Previous research has found that YP improves abnormal lipid metabolism in both animal models and in patients with T2DM [19]. However, the impact of YP on GDM and its underlying mechanism remains unclear.
In the present study, a high-fat and fructose diet (HFD)-induced GDM mouse model was employed to evaluate the effect of YP on GDM. Furthermore, changes in GM were assessed to explore the potential mechanism through which YP may exert therapeutic effects in GDM.
Six-week-old C57BL/6 mice were purchased from the Guangdong Medical Laboratory
Animal Center (Guangzhou, Guangdong, China). All animal experiments were approved
by the Ethics Committee of the First Affiliated Hospital of Guangdong
Pharmaceutical University. The female mice were randomly divided into three
groups: (1) the control group, which received a normal diet (10 kcal% fat, 3.85
kcal/g; Guangdong Medical Laboratory Animal Center; batch No. D12450B) and
untreated drinking water; (2) the model group, which received a high-fat diet (45
kcal% fat, 4.73 kcal/g; Guangdong Medical Laboratory Animal Center; batch No.
D12450B) and drinking water supplemented with 10% (w/v) fructose (Fru, 4 kcal/g;
Yongjin Biotechnology Co., Guangzhou, Guangdong, China; batch No. BWJ4341-2016).
This group was also named the HFD group; and (3) the treatment group (HFD + YP),
which received a daily oral gavage of YP powder (Shanghai Yuanye Bio-Technology
Co., Shanghai, China; batch No. S24912) dissolved in sterile water and
administered while maintained on an HFD, starting after mating. The dosage of YP
was 200 mg/kg body weight per day. The control group and HFD groups were given an
equal amount of sterile water by gavage after mating. After a six-week dietary
intervention, female mice were mated with lean male mice. Mating was confirmed by
the presence of a vaginal mucus plug the following morning, which was designated
as gestational day (GD) 0. Based on pilot experiments and power analysis, each
group required n = 6 mice to achieve 80% power to detect a 30% difference at
significance levels of
Mice underwent an oral glucose tolerance test (OGTT) at GD15 after a 12-hour fast. Each mouse was orally gavaged with glucose at a dose of 2 g/kg body weight, prepared in normal saline. Blood samples were collected from the vein at 0, 30, 60, 90, and 120 minutes after glucose administration to measure the glucose concentration.
Blood glucose levels were quantified using a glucose assay kit (Solarbio,
Beijing, China; batch No. BC2495) in accordance with the manufacturer’s protocol,
which follows the glucose oxidase method. Serum insulin levels were measured with
a mouse ultrasensitive insulin ELISA kit (Wuhan Colorful Gene Biotech Co., Ltd.,
Wuhan, China; batch No. JYM0351Mo). The HOMA-IR was calculated using the
following formula: HOMA-IR = (fasting glucose
3T3-L1 murine preadipocytes were purchased from Cas9X Biotechnology Company
(Suzhou, Jiangsu, China; batch No. TCM-C702). This cell line has been
authenticated by short tandem repeat analysis, morphological observation, and
adipogenic induction, and it tested negative for mycoplasma contamination.
3T3-L1 cells in the logarithmic growth phase were seeded into culture vessels at a
density of 2.0
Fecal samples were collected at GD16 in individual sterilized cages and immediately frozen in liquid nitrogen. DNA was extracted according to the instructions of the DNA extraction kits (TaKaRa Bio, Beijing, China; batch No. 9765). The integrity and purity of the DNA were detected by 1% agarose gel electrophoresis, and the concentration and purity of the DNA were detected by the NanoDrop One microvolume spectrophotometer (ThermoFisher Scientific, Waltham, MA, USA). PCR amplification and product electrophoresis were performed using genomic DNA as a template, and primers with barcodes and Premix Tag (TaKaRa Bio; Beijing, China; batch No. RR004A) were used for PCR amplification based on selected sequencing regions. Gene Tools Analysis Software (Cambridge, UK; Version 4.03.05.0, SynGene) was used to quantify PCR product concentrations. The volume of each sample was adjusted to ensure equal mass, and the PCR products were pooled. Mixed PCR products were purified using the E.Z.N.A. Gel Extraction and Gel Recovery Kit (Omega Bio-Tek, Guangzhou, Guangdong, China; batch No. D2500). Target DNA fragments were eluted with TE buffer. Library construction was then performed using the NEBNext® Ultra™ DNA Library Prep Kit for Illumina® (New England Biolabs, Ipswich, MA, USA; batch No. E7645) standard process. The high-throughput sequencing platform HiSeq was utilized for online sequencing, and the raw image data file was converted into a raw sequencing dataset (Raw Reads) through base recognition (Base Calling) analysis. The results were stored in the FASTQ file format, which contains the sequencing sequence (Reads) and its corresponding quality information. Alpha diversity, which reflects the richness and evenness of microbial communities within a sample, was evaluated using the Shannon and Simpson indices. Beta diversity, which measures compositional differences between microbial communities across samples, was evaluated using Principal Coordinates Analysis (PCoA) based on the Bray-Curtis distance matrix. Additionally, Analysis of Similarities (ANOSIM) was employed to determine the significance of differences in microbial communities between groups. Analysis of species differences across multiple groups was based on Kruskal-Wallis rank sum test. The false discovery rate (FDR) was used for multiple-testing correction.
The data are presented as mean
The experimental protocol for the animal study is shown in Fig. 1A. The HFD
intervention was initiated six weeks before pregnancy and was maintained
throughout the pregnancy. Fasting blood glucose was measured, and an OGTT was
performed before mating to exclude individuals with pre-existing diabetes. All
variables of interest satisfied the assumption of normality according to the
Shapiro-Wilk test (all p
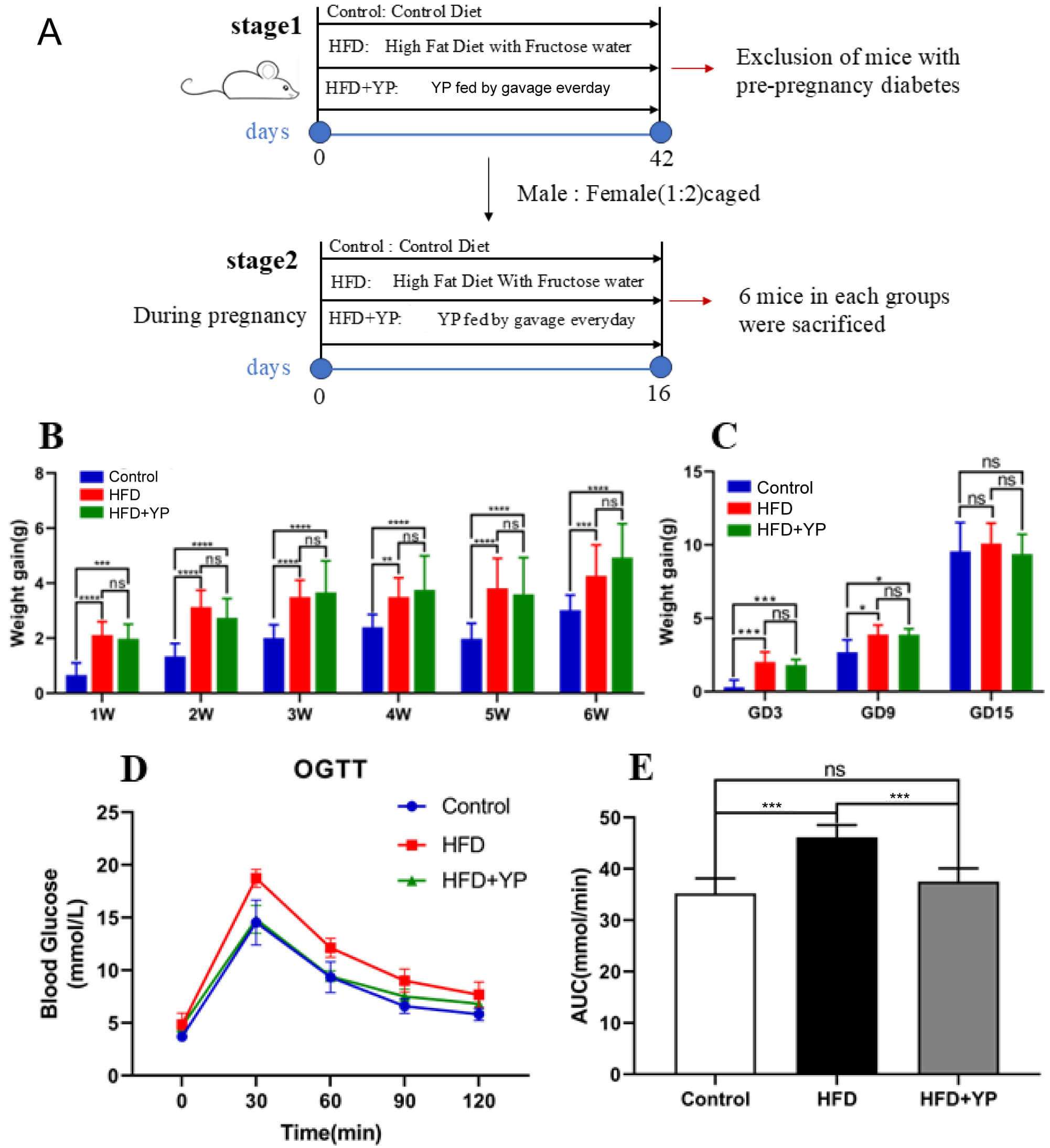 Fig. 1.
Fig. 1.
YP inhibits the progression of GDM. (A) Schematic design of the
experiment. (B) Weight gain during intervention before pregnancy. (C) Weight gain
during pregnancy. (D) Oral glucose tolerance tests (OGTT). (E) AUC of OGTT. *
p
Compared with the HFD group, the YP-treated group exhibited significantly lower
levels of blood glucose at 30 min, 60 min, and 90 min during the OGTT, along with
a smaller AUC (Fig. 1D,E). A notable decrease in HOMA-IR was also observed in the
HFD + YP group compared to the HFD group (1.00
Additionally, an ex vivo experiment was performed to evaluate the impact of YP on insulin resistance. Murine preadipocyte 3T3-L1 were induced to differentiate into adipocytes (Fig. 2A). Dexamethasone inhibited glucose uptake in adipogenically induced 3T3-L1 cells, indicating successful establishment of an insulin resistance model (Fig. 2B). YP was found to alleviate dexamethasone-induced impairment of glucose uptake in a dose-and time-dependent manner (Fig. 2C,D).
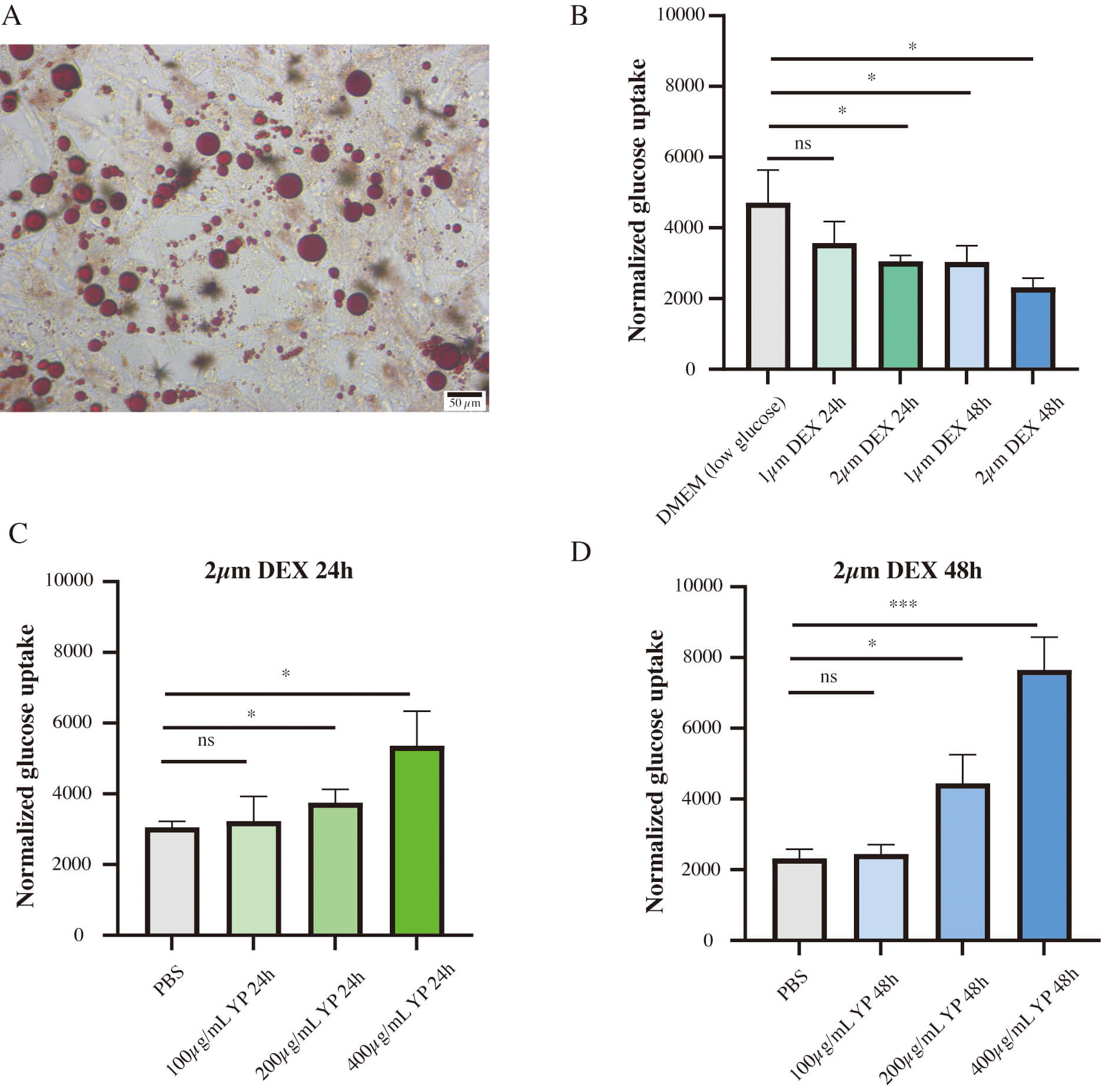 Fig. 2.
Fig. 2.
YP alleviated dexamethasone (DEX)-induced impaired glucose
uptake in 3T3-L1 adipocytes. (A) Red oil O staining demonstrated the
adipogenesis induction in 3T3-L1 cells, scale bar: 50 µm. (B) DEX inhibited
glucose uptake in 3T3-L1 adipocytes compared to control group. (C,D) YP enhanced
glucose uptake in 3T3-L1 adipocytes incubated with 2 µm DEX compared
to control group. ns, not significant; DMEM, Dulbecco’s Modified Eagle Medium; PBS, phosphate buffered saline, * p
Analysis of alpha diversity of GM among the three groups showed that the Shannon index was significantly higher in the HFD group than in the control group, whereas the Simpson index was lower. These findings indicated that the HFD-induced GDM model exhibited increased richness and decreased evenness of microbial communities compared with the control group. (Fig. 3A,B). In addition, YP prevented the HFD-induced increase in alpha diversity (Fig. 3A,B). Subsequently, the beta diversity of microbial communities among the three groups was investigated using PCoA, which revealed three distinct clusters corresponding to the intervention groups. The separation between the HFD and control groups was particularly pronounced, indicating HFD-induced alteration in GM composition. Importantly, YP administration induced additional alterations in the microbial community structure across samples (Fig. 3C).
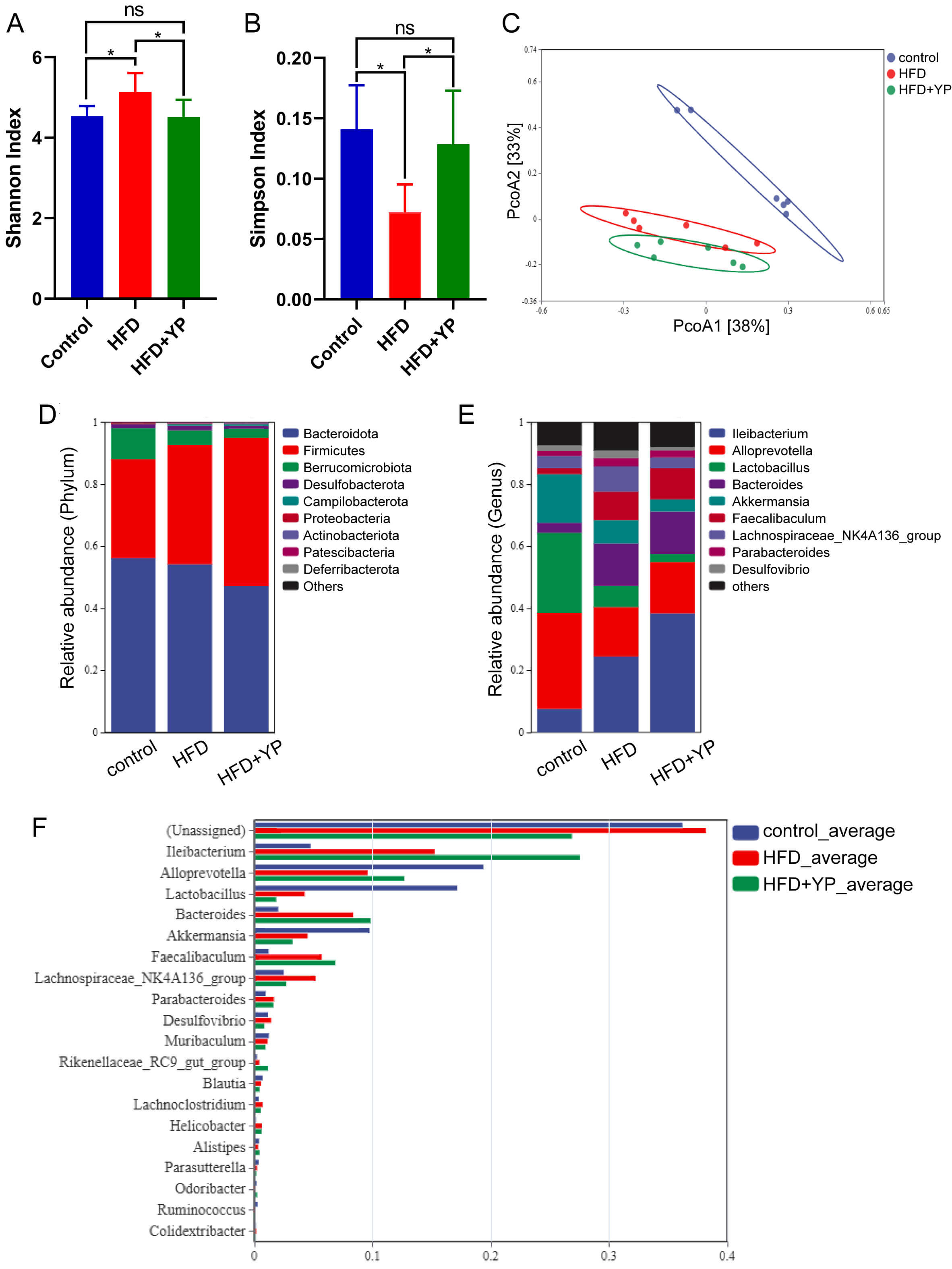 Fig. 3.
Fig. 3.
YP changed the composition of gut microbiota (GM). (A) Shannon
index in different groups. (B) Simpson index in different groups. (C) PCoA
analysis. (D) Relative abundance of microbial species of the top 10 phyla with
significant differences. (E) Relative abundance of microbial species of the top
10 genera with significant differences. (F) Relative abundance of microbial
species of the top 20 genera with significant differences. ns, no significance. *
p
We analyzed the relative abundance of species across the three groups at the phylum level (Fig. 3D). The Firmicutes/Bacteroidetes (F/B) ratio was increased in both the HFD and HFD + YP groups compared with the control group (Fig. 3D). An increased F/B ratio is commonly observed in obese individuals [20]. The elevated F/B ratio in the HFD and HFD + YP groups indirectly reflects the effect of the metabolic alteration induced by HFD treatment and is consistent with the finding that YP did not attenuate the HFD-induced weight gain in pregnant mice. At the genus level, the relative abundance of Alloprevotella decreases in the HFD group compared with the control group but increases in the HFD + YP group. In contrast, the relative abundance of Lachnospiraceae NK4A136 group increased in the HFD group but declined with YP treatment (Fig. 3E,F). These findings suggest that Alloprevotella and Lachnospiraceae NK4A136 group may be potential microbial targets through which YP alleviates GM dysbiosis and abnormal glucose metabolism.
This study investigated the effects of YP on GM in a GDM mouse model induced by an HFD. Although previous studies have reported the hypoglycemic effects of YP, its potential to improve glucose tolerance by modulating GM has not been explored. In this study, we defined the conventional GDM mouse models, which were typically induced by either HFD or high-fructose intake alone. This refinement is particularly relevant, as modern diets often combine high levels of both fat and fructose-nutritional factors closely linked to obesity, a key contributing factor for GDM [21, 22].
In this study, we found that HFD treatment impaired glucose tolerance,
establishing a reliable model for GDM. YP demonstrated a significant effect in
improving glucose tolerance, likely by enhancing insulin secretion. Furthermore,
YP treatment increased the abundance of Alloprevotella, while decreasing
the abundance of the Lachnospiraceae NK4A136 group. Growing evidence
links systemic inflammation with insulin resistance, a key feature of all types
of diabetes, including GDM. An HFD can increase the abundance of
lipopolysaccharide (LPS)-producing gut bacteria, which in turn raises blood
glucose levels and triggers systemic inflammation [23, 24]. Pregnant mice
subjected to a high-fructose diet exhibited increased weight gain, elevated
fasting glucose, and insulin resistance through the activation of the
NF-
The Lachnospiraceae NK4A136 group was found to be enriched during early
pregnancy among women who were later diagnosed with GDM compared with the healthy
controls [33]. Additionally, an increased abundance of Lachnospiraceae
NK4A136 group was observed in a rat model of advanced-stage type 1 diabetes [34].
The Lachnospiraceae NK4A136 group was positively correlated with tumor
necrosis factor-alpha (TNF-
The study has several limitations. Firstly, YP administration was evaluated at a single dose, preventing any assessment of potential dose-dependent therapeutic effects on GDM. Secondly, previous research has shown that HFD can induce placental oxidative stress and vascular dysregulation in pregnant mice [39]. For example, Sanches et al. [40] reported that HFD feeding led to fetal growth restriction by altering placental thickness and nutrient transport. However, the effects of YP on placental development and fetal growth have not yet been evaluated, and these aspects will be addressed in future studies.
YP has the potential to improve glucose intolerance in GDM mice induced by HFD. The regulation of GM is one of the mechanisms underlying the antidiabetic effect of YP in GDM. These findings provide novel insights into the biological activities of YP in the treatment of GDM.
The data presented in this study is available from the corresponding author on reasonable request.
PJ and MH designed the research study. SZ and ZD performed the research. BX, XH and WX analyzed the data. All authors contributed to critical revision of the manuscript for important intellectual content. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
All animal experiments meet “3Rs” principle. All animal experiments were performed in compliance with the Guide of the Ethics Committee of the First Clinical Medicine School of Guangdong Pharmaceutical University. The ethics approval number is 821015321.
We would like to express our gratitude to Dr. De-pei Li from Sun Yat-sen University who helped us during the writing and revising of this manuscript.
The study was funded by National Natural Science Foundation of China (82104910).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.