





1 Diagnostic Radiography Technology (DRT) Department, College of Nursing and Health Sciences, Jazan University, 45142 Jazan, Saudi Arabia
Abstract
Seasonal fluctuations in environmental conditions, particularly variations in photoperiod and ambient temperature, influence diverse physiological and behavioral processes. With advances in functional magnetic resonance imaging (fMRI), researchers have begun to investigate how such environmental factors modulate intrinsic brain activity. This review synthesizes current evidence on how seasonal, circadian, and environmental variations affect brain functional connectivity.
A comprehensive search of PubMed/Medical Literature Analysis and Retrieval System Online (MEDLINE), Scopus, and Web of Science databases was conducted for peer-reviewed fMRI studies published between 2010 and 2025. Eligible studies investigated the impact of environmental factors, such as photoperiod, temperature, and time of day, on brain functional connectivity. Due to methodological heterogeneity, a narrative synthesis approach was used in accordance with Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) 2020 guidelines. The review process is illustrated using a PRISMA flow diagram, and a checklist is provided as supplementary material.
Sixteen original studies spanning cross-sectional and longitudinal designs met the inclusion criteria. Seasonal and environmental exposures modulated connectivity in major networks, particularly the default mode network (DMN), salience network (SN), and sensorimotor network (SMN). For the DMN and SMN, photoperiod consistently affected connectivity, with peaks in autumn and reductions in summer. Circadian and temperature effects were also observed. Methodological factors, including scanner stability, emerged as critical.
Environmental rhythms significantly affect brain connectivity and should be considered in neuroimaging research and clinical practice. These effects are sometimes comparable to clinical conditions. Future work should enhance control of environmental exposures and incorporate physiological markers to improve reproducibility.
PROSPERO (CRD420261390073).
Keywords
- magnetic resonance imaging
- brain
- seasons
- circadian rhythm
- photoperiod
- neural pathways
- default mode network
Seasonal changes in environmental conditions, particularly variations in daylight length (photoperiod), temperature, and light exposure, have long been found to influence human physiology, behavior, and mental health. These changes impact mood, sleep, cognition, and biological rhythms throughout the year [1, 2]. Clinically, seasonal fluctuations are most prominently observed in seasonal affective disorder (SAD), a subtype of depression that affects 1–10% of the population in temperate regions and is characterized by recurrent depressive episodes during fall and winter, typically triggered by reduced daylight exposure [3, 4]. However, subclinical seasonal variations in mood, sleep patterns, cognitive performance, and social behavior occur across the general population, suggesting that seasonal environmental changes exert broad influences on brain function [5].
Human brain function is affected by intrinsic neurobiological mechanisms as well as external environmental influences. Among these, seasonal variations in photoperiod, ambient temperature, and climatic patterns are increasingly recognized as important modulators of neural activity and behavior [2, 6]. The neurobiological substrates of these seasonal effects are being explored increasingly often through neuroimaging techniques, particularly functional magnetic resonance imaging (fMRI), which has allowed researchers to examine how seasonal and environmental factors modulate brain activity (Fig. 1). fMRI has revolutionized our understanding of brain function by enabling the non-invasive measurement of neural activity through blood-oxygen-level-dependent (BOLD) signals [7]. Functional connectivity is defined as the temporal dependency of neuronal activation patterns of anatomically separated brain regions. In recent years, an increasing body of neuroimaging studies has explored functional connectivity by measuring the level of co-activation of resting-state fMRI (rs-fMRI) time-series between brain regions [8]. rs-fMRI enables the study of intrinsic functional connectivity so that the temporal synchronization of neural activity between anatomically distinct brain regions can be captured while the individual is not engaged in a task. rs-fMRI measures spontaneous fluctuations in BOLD signals, which enables the mapping of intrinsic brain networks such as the default mode network (DMN), sensorimotor network (SMN), and salience network (SN) [7, 8]. These networks are critical for fundamental functions, including emotion regulation, cognition, and self-referential thought.
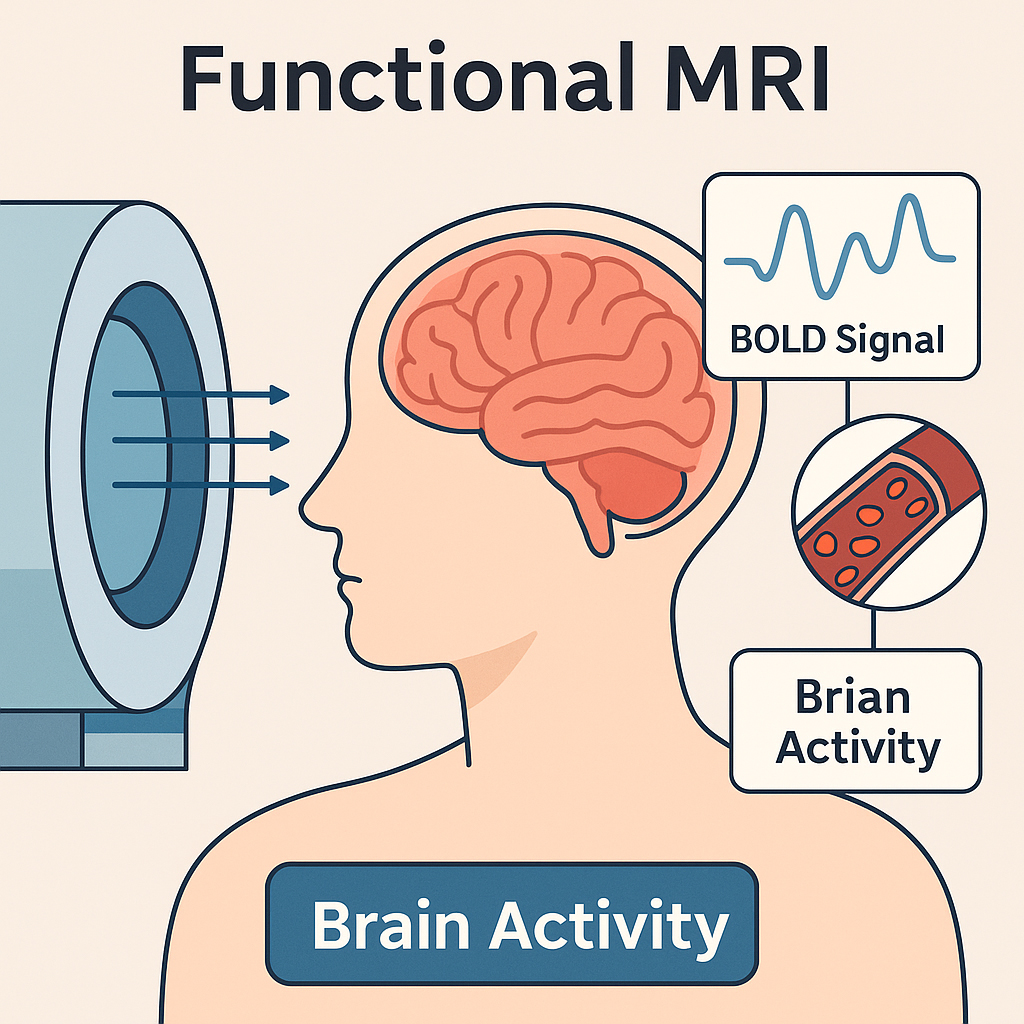 Fig. 1.
Fig. 1.
Functional magnetic resonance imaging (fMRI) methodology for detecting brain activity. fMRI uses the blood-oxygen-level-dependent (BOLD) signal to non-invasively detect brain activity. Neural activation increases local blood flow, altering oxygenation levels that are captured through high-field magnetic resonance imaging (MRI) scanners. This schematic diagram was created by the author using Adobe Illustrator 2024 (Adobe Inc., San Jose, CA, USA) and BioRender.com (https://www.biorender.com/). The author has no financial or personal relationship with BioRender.com, and the use of this tool does not imply any endorsement.
Recent studies suggest that functional connectivity within and between these networks shows seasonal modulation. Studies have shown seasonal changes in brain metabolism [6], functional network strength [9], and regional activation patterns, especially in regions associated with affect regulation, sensory processing, and memory [10]. A previous study reported robust seasonal variation in both global and regional functional connectivity, with peak connectivity in autumn and the lowest levels in summer [9]. Changes were observed in the DMN, SMN, and ventral attention network (VAN), as well as in white matter (WM) connectivity metrics such as network density and efficiency. Similarly, Zhang et al. [11] and Triana et al. [12] identified strong seasonal patterns in the SMN over a multi-year single-subject dataset, linked to photoperiod-related molecular changes. These results highlight a reproducible seasonal signature in functional neural network.
Moreover, environmental factors such as temperature and daylight length have been implicated as modulators of neural activity. Granés et al. [10] found that higher ambient temperatures were associated with reduced functional connectivity in the medial parietal, hippocampal, and SN among preadolescents. These environmental effects on brain function are not merely incidental, and they may have critical developmental, behavioral, and clinical implications.
The neurobiological pathways through which seasonal environmental changes influence brain function involve multiple interconnected systems. Photoperiod variations affect the suprachiasmatic nucleus, the brain’s master circadian clock, which regulates melatonin secretion and downstream hormonal cascades [2]. Temperature changes influence peripheral and central thermoreceptors, affecting hypothalamic regulatory centers and their connections to cortical and limbic regions [6]. These environmental inputs converge on brain networks critical for mood regulation, attention, and social cognition. The DMN, comprising the medial prefrontal cortex, posterior cingulate cortex, and angular gyrus, serves as a hub for self-referential processing and has been consistently implicated in mood disorders [8]. Similarly, the SN, anchored by the anterior insula and dorsal anterior cingulate cortex, plays crucial roles in attention and emotional processing [7].
However, not all studies support the presence of true biological seasonal effects. Di et al. [13] cautioned that scanner-related factors such as signal drift might mimic seasonal variation, emphasizing the need for methodological rigor and scanner quality control in such analyses. Despite these challenges, understanding the seasonality of brain function has important implications for clinical neuroimaging. The magnitude of seasonal effects observed in functional connectivity is comparable to that seen in several neurological and psychiatric conditions [9], raising concerns about the potential for confounding effects in clinical trials or diagnostic imaging conducted across different seasons.
Therefore, this review aims to synthesize current evidence concerning seasonal effects on brain functional connectivity as measured by fMRI. Specifically, this study (1) identifies which brain networks exhibit seasonal variation, (2) examines relationships between environmental variables and neural connectivity, (3) assesses the methodological strengths and limitations of existing studies, and (4) explores implications for both clinical and research settings in neuroscience. Despite its neuroscientific focus, this review holds considerable relevance for hospital-based clinical practice. Seasonal modulation of brain connectivity may influence neuroimaging interpretation, treatment timing, and research design within hospital settings, particularly in radiology, psychiatry, and neurology departments. By addressing its aims, this review seeks to establish seasonal neuroscience as a critical consideration in both basic and clinical neuroscience while providing a roadmap for future investigations in this rapidly evolving field.
This review followed Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) 2020 guidelines, and a checklist is provided as supplementary material (Supplementary File 1). The systematic review was registered in the Internationa Prospective Register of Systematic Reviews (PROSPERO) database (CRD420261390073, https://www.crd.york.ac.uk/PROSPERO/view/CRD420261390073). This comprehensive review employed a systematic approach to identify and synthesize studies examining seasonal effects on brain functional connectivity (Fig. 2). The search strategy was designed to encompass the multidisciplinary scope of this emerging research area, integrating terminology from neuroscience, environmental science, clinical psychiatry, and related fields. Multiple databases were thoroughly searched to ensure extensive coverage. These included primary databases such as PubMed/Medical Literature Analysis and Retrieval System Online (MEDLINE) (https://pubmed.ncbi.nlm.nih.gov/), Scopus (https://www.scopus.com/), and Web of Science (https://www.webofscience.com/). The search utilized a detailed and structured strategy, combining medical subject headings (MeSH) terms and free-text keywords with Boolean operators. The primary search string was constructed as follows: (“seasonal” OR “photoperiod” OR “circadian” OR “daylight” OR “temperature” OR “climate”) AND (“brain” OR “neural” OR “neuroimaging”) AND (“functional connectivity” OR “resting state” OR “fMRI” OR “network” OR “BOLD”) AND (“seasonal affective” OR “environmental neuroscience” OR “seasonal variation”). This comprehensive and precise search strategy facilitated the identification of relevant articles, ensuring that critical studies spanning diverse, yet interconnected disciplines were considered.
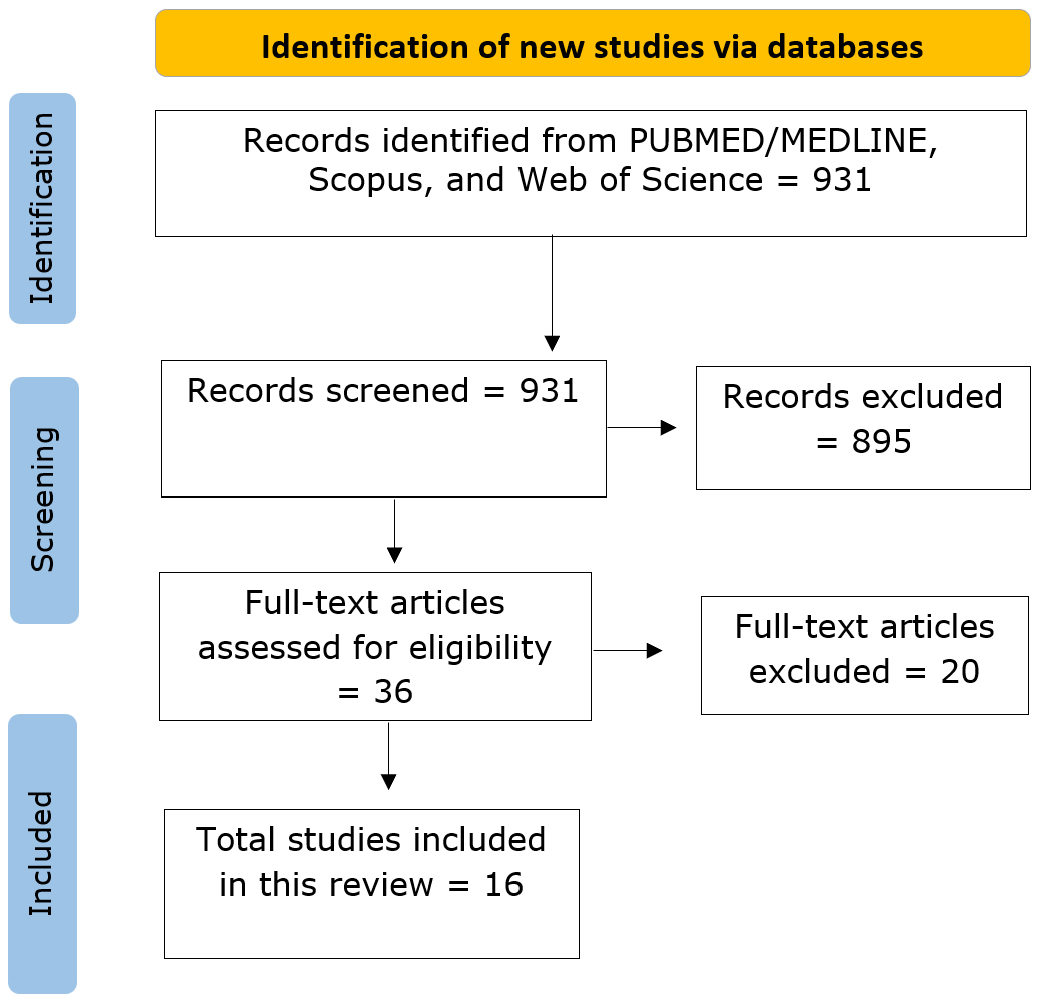 Fig. 2.
Fig. 2.
Flowchart depicting the systematic literature search and study selection process. Figure created by the author using Microsoft Visio (Microsoft Corporation, Redmond, WA, USA, https://www.microsoft.com/en-us/microsoft-365/visio/flowchart-software). Abbreviation: MEDLINE, Medical Literature Analysis and Retrieval System Online.
Studies were included if they met all of the following criteria: (1) published in peer-reviewed journals between 2010 and 2025; (2) employed fMRI, including rs-fMRI or task-based paradigms; (3) investigated the effects of seasonal, circadian, or environmental variables, such as photoperiod, temperature, or time of day, on brain functional connectivity; and (4) involved human participants with clearly reported sample sizes. Additional exclusion criteria were applied to refine the study pool by eliminating studies that, while meeting some inclusion parameters, lacked relevance or methodological appropriateness. Specifically, studies were excluded if they: (1) were not published in English, (2) were conducted solely on animals or in vitro models, (3) lacked functional neuroimaging data (e.g., used only structural magnetic resonance imaging (MRI) or behavioral outcomes), (4) were single-subject or case reports, or (5) did not include any environmental or temporal variables related to seasonality.
Due to the heterogeneity of imaging methodologies, seasonal variables, and outcome measures across the included studies, a quantitative meta-analysis was not feasible. Therefore, a narrative synthesis approach was employed to integrate findings across studies.
For each included study, the following information was systematically extracted: study characteristics (authors, year, country, study design), sample characteristics (sample size, age range, gender distribution, clinical status), environmental variables (photoperiod, temperature, season definitions), neuroimaging parameters (scanner type, field strength, acquisition parameters), analysis method (connectivity metrics), and key findings (affected brain networks, seasonal patterns).
The reviewed studies demonstrate that seasonal and environmental factors exert significant modulatory effects on brain functional connectivity across diverse populations and methodological frameworks. These effects emerge from both rs-fMRI and task-based fMRI paradigms, encompassing daylight duration, ambient temperature, time of day, and environmental stimuli exposure.
The comprehensive literature search identified 931 potentially relevant records across all databases. After removing duplicates and applying inclusion/exclusion criteria, 16 studies met the final inclusion criteria for this review. These studies were published between 2011 and 2025, with a notable increase in publications after 2020, reflecting growing interest in environmental neuroscience. Table 1 (Ref. [6, 9, 10, 11, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24]) presents a comprehensive summary of the study characteristics included in this review. The review combines findings from all or most available fMRI studies that examined the influence of seasonal, circadian, and environmental factors on brain functional connectivity. These studies span a broad range of populations, from children to older adults, and were conducted across diverse geographic regions, including North America, Europe, and Asia. Both cross-sectional and longitudinal designs were represented, with some studies employing dense sampling or test-retest paradigms to capture intraindividual variability. Sample sizes varied substantially, ranging from small, intensively studied cohorts to large-scale population datasets. Participant demographics also differed, with most studies including mixed-gender groups, while others focused on specific populations such as athletes. Age ranges cover childhood through late adulthood, enhancing the generalizability of findings across the lifespan. All studies employed high-field MRI systems, primarily 3T (Tesla) scanners, and most used rs-fMRI to examine intrinsic brain network dynamics. A minority used task-based fMRI to probe cognitive and emotional processes under seasonal influence. Environmental exposures included seasonal timing, photoperiod, ambient temperature, and time of day, with some studies spanning multiple time points to assess longitudinal changes. A few investigations also addressed potential confounds such as scanner variability. Brain networks frequently examined included the DMN, SN, SMN, visual network (VN), and attention network (AN); key limbic regions were also commonly examined. Analytic approaches ranged from seed-based connectivity to graph theory metrics and independent component analysis. Together, these studies provide a robust foundation for understanding how environmental rhythms modulate functional neural network.
| Study authors | Country | Design | Sample size | Age range | Gender | Scanner strength | fMRI type | Seasonal/Circadian variable | Main brain regions | Key findings |
| Alyssia Wilson et al. [14] | Canada | Longitudinal pre-post season | 19 female athletes (12 collision, 7 contact) | 18–21 years | Female | 3T | rs-fMRI | Athletic season (pre vs post, 166 days apart) | DMN: Superior parietal lobule, supramarginal gyrus, superior frontal gyrus, frontal pole; DAN: Superior frontal gyrus, supramarginal gyrus | Decreased DMN connectivity over season (contact group); Increased DAN connectivity in collision vs contact athletes; Potential cumulative effects of repetitive head impacts on brain connectivity |
| Ahmed Abou Elseoud et al. [15] | Finland | Cross-sectional case-control | 45 SAD patients, 45 controls | 39.78 |
Mixed (30 F, 15 M in SAD group) | 1.5T | rs-fMRI | Winter seasonal affective disorder | Visual networks (RSNs 5–11), sensorimotor networks (RSNs 1–3), attention network (RSN 4) | SAD patients showed increased functional connectivity in 11/47 RSNs; higher ALFF in visual and right sensorimotor cortex; increased connectivity in visual, sensorimotor, and attentional networks |
| CONNOR W. GHILES et al. [16] | USA | Longitudinal cohort | 17 male football players | 15–17 years (mean 16 |
Male | 3T | rs-fMRI | Single football season (pre vs post) | Frontal lobe, parietal lobe, mediotemporal networks | Decreased local efficiency (–24.9%) and strength (–14.5%) in frontal network; decreased strength (–7.5%) in parietal network; increased efficiency (+55.0%) and strength (+47.4%) in mediotemporal networks |
| Xin Di et al. [13] | Multiple (USA, Germany) | Longitudinal multi-dataset | 8 individuals | 23–45 years | Mixed | 3T | rs-fMRI | Weather parameters (daylight, temperature, etc.) | None (effects attributed to scanner artifacts) | Weather parameters could be predicted from brain data, but similar predictions achieved with anatomical images and phantom scans, suggesting scanner stability effects rather than brain function changes |
| Laura Granés et al. [10] | Netherlands | Cross-sectional | 2229 children | 9–12 years (mean 10.1 |
Mixed (51.5% female) | 3T | rs-fMRI | Daily ambient temperature (7-day period before scan) | Medial parietal network, salience network, hippocampus | Higher temperatures (17.7–25.5 °C) associated with lower within-network connectivity in medial parietal, salience networks, and hippocampus; effects strongest day before scan |
| Jeanne Racicot et al. [17] | Canada | Longitudinal | 6 healthy participants | 24–32 years | 5 females, 1 male | 3T Siemens Magnetom Trio | rs-fMRI | Day-to-day fluctuations over days, weeks, months (40–50 scans per participant) | Limbic areas (amygdala, hippocampus, insula), prefrontal cortex | Limbic functional connectivity explained 15% and 11% of within-person variation in positive/negative affect; predicted ~5% and 2% of new data variation |
| Bradley Fitzgerald et al. [18] | USA | Longitudinal | 42 football athletes, 14 controls | Football: 16.7 |
Male only | 3T General Electric Signa HDx | rs-fMRI | Seasonal across football season (pre-season, 2 in-season, post-season) | Whole-brain (248 regions), seven functional networks, somatomotor and ventral attention networks | Decreased FC self-similarity during late season with post-season recovery; correlated with cumulative head impacts; somatomotor/ventral attention most affected |
| Derek C Monroe et al. [19] | USA | Longitudinal | 24 athletes (13 soccer, 11 controls) | 20.2 |
Male NCAA Division I | 3T Philips Achieva | rs-fMRI | Seasonal across soccer season (117.3 |
Central autonomic network (periaqueductal grey, hypothalamus, amygdala, ventral anterior insula, medial prefrontal/orbitofrontal cortex) | Head-to-ball impacts increased FC in autonomic network; left insula-orbitofrontal connectivity linked to diminished proactive cognitive control |
| Lyuan Xu et al. [9] | USA | Cross-sectional | 410 | 26–35 years | 247 F, 163 M | 3T | rs-fMRI | Seasonal variations (Winter, Spring, Summer, Autumn) | GM and WM, 14 large-scale networks, sensorimotor, posterior insula, ventral DMN | Seasonal variations in FC with autumn highest, summer lowest. Environmental factors (temperature, day length) significantly associated with brain activity |
| Christelle Meyer et al. [6] | Belgium | Cross-sectional | 28 | 21 |
14 F, 14 M | 3T | Task-based fMRI | Seasonal timing across different seasons | Thalamus, amygdala, frontal areas, hippocampus, sensorimotor, posterior insula, ventral DMN | Task-specific seasonal patterns: attention peaked at summer solstice, working memory peaked at autumn equinox |
| Csaba Orban et al. [20] | Singapore | Cross-sectional with test-retest | 942 (S1), 869 (S2) | Young adults | 60% female | 3T | rs-fMRI | Time of day (8 AM–10 PM) | Global signal, sensory-motor, visual cortex, somatomotor, anterior cingulate, putamen, thalamus | Paradoxical reduction in global signal fluctuation from morning to evening; widespread decreases in FC throughout day |
| Allen A Champagne et al. [21] | Canada | Longitudinal | 23 collegiate football players | 20–21 years | Male | 3T Siemens | rs-fMRI with dual-echo pCASL | Not applicable (sub-concussive impacts focus) | DMN | DMN hyper-connectivity throughout season, independent of cerebrovascular physiology changes |
| Rui Zhang et al. [11] | USA | Longitudinal single-subject | 2 subjects (MyConnectome: 84 sessions; Kirby: 156 sessions) | 40–45 years | Male | 3T Siemens/ Philips | rs-fMRI | Day length and day-to-day variations | Sensorimotor network, visual network | Day length associated with increased sensorimotor network connectivity; caffeine enhanced seasonal effects |
| Linfeng Hu et al. [22] | USA | Cross-sectional analysis (ABCD) | 4102 | ~10 years (median 120 months) | 53.1% female | 3T | rs-fMRI | Time of day, time of week, time of year | Visual, dorsal attention, salience, frontoparietal, basal ganglia, DMN, somatomotor | Later scan times |
| Simone Kühn et al. [23] | Germany | Experimental | 24 | Mean 28.5 years | 16 F, 8 M | 3T | rs-fMRI during passive viewing | Environmental context (natural vs built) | DMN, DAN, VAN, somatomotor | Higher FC for natural vs built environments in DAN-VAN, DAN-DMN, DMN-somatomotor; urban upbringing negatively correlated with natural connectivity |
| Liucija Vaisvilaite et al. [24] | Norway | Cross-sectional analysis (HCP) | 594 | 22–35 years | 310 F, 284 M | 3T | rs-fMRI | Time of day (6 timespans: 09:00–21:00) | DMN, CEN, SN | No changes in effective connectivity across time of day, but hemodynamic parameters (BOLD signal) showed significant diurnal effects; decay/amplitude reduced in morning (9–13 h) |
Abbreviations: ABCD, Adolescent Brain Cognitive Development; ALFF, Amplitude of Low-Frequency Fluctuation; CEN, Central Executive Network; DAN, dorsal attention network; DMN, default mode network; F, Female; FC, functional connectivity; fMRI, functional magnetic resonance imaging; rs-fMRI, resting-state fMRI; GM, gray matter; HCP, Human Connectome Project; M, Male; NCAA, National Collegiate Athletic Association; pCASL, pseudo-Continuous Arterial Spin Labeling; RSN, Resting State Network; S1, primary somatosensory cortex; SAD, seasonal affective disorder; SN, salience network; T, Tesla; VAN, ventral attention network; WM, white matter.
The comprehensive analysis of study characteristics reveals several important patterns in environmental neuroscience research (Table 2, Ref. [6, 9, 10, 11, 13, 14, 15, 16, 17, 19, 20, 22, 23, 24]). The field shows methodological diversity with balanced representation of cross-sectional and longitudinal designs, though a notable geographic bias toward North American and European populations limits global generalizability. The predominant focus on DMN (75% of studies) reflects its clinical relevance in mood and cognitive disorders commonly seen in hospital settings. The equal distribution across sample size categories (small, medium, large) reinforces confidence in findings across different scales of investigation. Notably, the inclusion of both healthy and clinical populations, along with specialized athletic cohorts, demonstrates the broad applicability of seasonal effects across diverse patient populations encountered in hospital medicine.
| Study classification | Number of studies | Percentage | Examples |
| Original research studies | 16 | 100% | All included studies |
| Review studies | 0 | 0% | Excluded by criteria |
| Adult studies ( |
12 | 75% | [6, 9, 20] |
| Pediatric studies ( |
3 | 19% | [10, 16, 22] |
| Mixed age range | 1 | 6% | [24] |
| Photoperiod/Daylight | 8 | 50% | [6, 9, 11] |
| Temperature | 3 | 19% | [10, 13] |
| Time of day (Circadian) | 4 | 25% | [20, 22, 24] |
| Seasonal timing | 6 | 38% | [6, 9, 15] |
| Athletic season effects | 5 | 31% | [14, 16, 19] |
| Multiple environmental factors | 2 | 13% | [13, 22] |
| Cross-sectional | 9 | 56% | [6, 9, 10] |
| Longitudinal | 7 | 44% | [11, 14, 17] |
| Small (20–50) | 6 | 38% | [6, 17, 23] |
| Medium (51–500) | 4 | 25% | [9, 15, 24] |
| Large ( |
6 | 38% | [10, 20, 22] |
| rs-fMRI | 15 | 94% | Majority of studies |
| Task-based fMRI | 1 | 6% | [6] |
| North America | 9 | 56% | USA and Canada |
| Europe | 5 | 31% | Finland, Netherlands, Belgium, Germany, Norway |
| Asia | 2 | 13% | Singapore |
| DMN | 12 | 75% | Most frequently studied |
| SMN | 10 | 63% | Second most common |
| AN | 8 | 50% | Dorsal and ventral attention |
| SN | 6 | 38% | Temperature-sensitive |
| VN | 5 | 31% | Circadian modulation |
| Healthy controls only | 11 | 69% | Typical environmental studies |
| Clinical populations | 1 | 6% | SAD patients [15] |
| Athletic populations | 4 | 25% | Sports-related studies |
Abbreviations: DMN, default mode network; SMN, sensorimotor network; AN, attention network; SN, salience network; VN, visual network; SAD, seasonal affective disorder.
Studies employed diverse approaches to characterize seasonal and environmental variables. Photoperiod was the most investigated factor, typically calculated using astronomical formulas based on study location and date. Systematic seasonal variations in brain connectivity patterns correlate with environmental variables, such as day length. In a previous study using a cross-sectional dataset of over 400 participants, Xu et al. [9] reported significant seasonal variations in gray matter (GM) and WM functional connectivity, with the DMN, sensorimotor network, and posterior insula network peaking in autumn and reaching minimum levels in summer. Meyer et al. [6] utilized task-based fMRI to demonstrate that attention-related activity peaked around the summer solstice, while working memory activations were highest during the autumn equinox, indicating photoperiodic influences on cognitive resource allocation. Dense longitudinal data spanning over 240 sessions across two subjects from Zhang et al. [11] revealed that longer day lengths correlated with increased SMN and VN connectivity, with effects amplified by caffeine intake. Triana et al. [12] identified seasonal fluctuations in sustained attention and cortical rhythms using combined magnetoencephalography (MEG) and fMRI across multiple months. The neurobiological basis involves neurochemical mechanisms, as Meyer et al. [6] found that serotonin transporter binding in prefrontal and limbic regions varied with daylight exposure, correlating with photoperiod-sensitive functional connectivity. Mc Mahon et al. [25] demonstrated that light-responsive neurotransmitter systems directly influence network organization. They identified seasonal patterns in cerebral metabolic rate of oxygen consumption, particularly in the precuneus, providing a mechanistic link between photoperiod changes and network organization. Clinical relevance is evident in research on SAD. Abou Elseoud et al. [15] found heightened connectivity in visual and attentional networks during winter in SAD patients compared to controls, interpreted as compensatory mechanisms for reduced light input, suggesting pathological seasonal changes reflect exaggerated responses within normal photoperiod-responsive systems.
Temperature was examined in several studies, using either direct meteorological measurements or regional weather data. Environmental temperature significantly modulates brain connectivity through acute and chronic mechanisms. Granés et al. [10] investigated over 2000 children aged 9–12 and revealed that higher ambient temperatures decreased connectivity in medial parietal networks, the hippocampus, and the SN. Effects were strongest for temperatures measured the day before scanning, indicating lagged neural responses to thermal changes. Mechanisms may involve thermoregulatory demands on neural resources, direct temperature effects on neural metabolism, or thermal-neurovascular coupling interactions [10]. Temperature-induced alterations in cerebral blood flow could influence BOLD signals underlying connectivity measurements, though specific thermal-neurovascular relationships require further investigation [7]. Di et al. [13] raised critical methodological concerns, demonstrating that weather parameters could be predicted based on fMRI data using anatomical scans and phantom data, suggesting that some reported seasonal effects might reflect scanner instabilities rather than neurobiological changes. This underscores the need for rigorous controls in environmental neuroscience. Temperature effects have climate change implications, as understanding thermal influences on neural function has become crucial for public health planning as global temperatures rise and extreme weather events increase [10].
Circadian timing critically influences connectivity patterns. Orban et al. [20] examined over 800 adults scanned between 8:00 AM and 10:00 PM, observing paradoxical reductions in global signal fluctuation and decreased SMN and VN connectivity as days progressed, suggesting circadian-driven changes alter baseline network dynamics. Hu et al. [22] found later daily scans associated with lower network efficiency, clustering, and stability in visual, dorsal attention, and salience networks. Weekend and summer scans showed distinct profiles, highlighting circadian-seasonal interactions in network organization. Racicot et al. [17] demonstrated functional significance through 40–50 scanning sessions per participant, finding limbic network connectivity variations (amygdala, hippocampus) explained significant within-subject variance in affective states, suggesting environmental context meaningfully shapes emotional brain architecture.
Seasonal changes in brain network connectivity are demonstrated in Fig. 3 below. Environmental factors (photoperiod, temperature, time of day) modulate brain functional connectivity, resulting in weaker (top) or stronger (bottom) inter-network coupling patterns. The arrows in the figure indicate environmental influences on neural network organization, demonstrating the dynamic relationship between external conditions and brain connectivity strength.
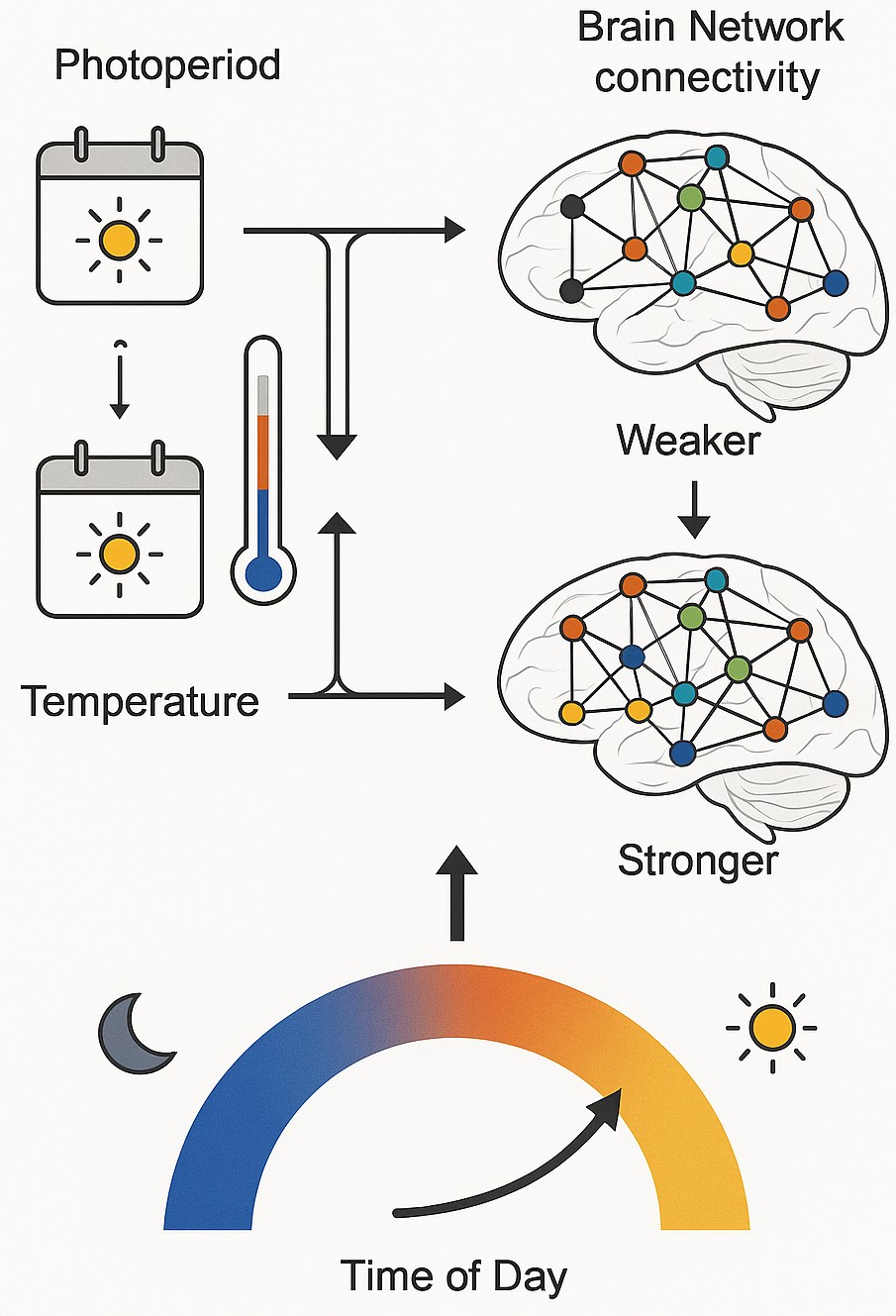 Fig. 3.
Fig. 3.
Seasonal changes in brain network connectivity. Arrows indicate environmental influences on neural network organization, demonstrating the dynamic relationship between external conditions and brain connectivity strength. This figure was created by the author using Adobe Illustrator 2024 (Adobe Inc., San Jose, CA, USA) and BioRender.com (https://www.biorender.com/).
Longitudinal athletic studies reveal activity-dependent neural plasticity. Wilson et al. [14] reported decreased DMN connectivity in female collision and contact sport athletes post-season, while Monroe et al. [19] found decreased DMN connectivity in male soccer players exposed to repetitive head impacts. Both studies observed increased dorsal attention network (DAN) connectivity, suggesting compensatory responses to subconcussive impacts. Ghiles et al. [16] found post-season reductions in local efficiency and connectivity in frontal and parietal regions of male high school football players, with mediotemporal network increases reflecting differential vulnerability and compensatory reorganization. Fitzgerald et al. [18] demonstrated decreased network self-similarity in football athletes, particularly in the somatomotor network and VAN, correlating with cumulative head impact exposure. Monroe et al. [19] identified increased central autonomic network connectivity in soccer players exposed to repetitive impacts, particularly in left insula-orbitofrontal pathways linked to impaired cognitive control. Champagne et al. [21] found DMN hyperconnectivity across collegiate football seasons using dual-echo pseudo-Continuous Arterial Spin Labeling (pCASL) fMRI, with changes unexplained by cerebrovascular factors, supporting functional mechanisms from subconcussive impacts.
The DMN demonstrates seasonal variations across multiple studies in this review. The DMN, comprising the medial prefrontal cortex, posterior cingulate cortex, precuneus, and angular gyrus, exhibits seasonal sensitivity that affects within-network integration and between-network interactions. Xu et al. [9] reported significant seasonal variations in ventral DMN connectivity, with significantly higher connectivity in autumn than in other seasons, supporting the notion that the DMN may be a driving factor for seasonal connectivity changes. Meyer et al. [6] found that working memory-related DMN activation peaked during the autumn equinox, indicating photoperiodic influences on cognitive resource allocation. The DMN’s role in self-referential processing, episodic memory, and future planning [8] makes its seasonal sensitivity functionally significant. Studies examining athletic populations also revealed DMN changes: Wilson et al. [14] reported decreased DMN connectivity in female athletes over a sports season, while Champagne et al. [21] found DMN hyperconnectivity throughout collegiate football seasons. Additional evidence comes from clinical populations, where Abou Elseoud et al. [15] demonstrated heightened DMN connectivity in seasonal affective disorder patients during winter, interpreted as compensatory mechanisms for reduced environmental input.
SMNs show seasonal modulation linked to photoperiod duration across multiple studies. These networks, encompassing primary motor cortex, supplementary motor areas, and somatosensory cortex, exhibit systematic fluctuations potentially reflecting adaptive changes in motor control and sensory processing. Xu et al. [9] found that sensorimotor network connectivity peaked in autumn and reached minimum levels in summer. Zhang et al. [11] provided dense longitudinal evidence spanning over 240 sessions across two subjects, revealing that longer day lengths correlated with increased SMN connectivity, with effects amplified by caffeine intake. This study demonstrated highly reproducible seasonal patterns closely aligned with photoperiod changes. Meyer et al. [6] observed that sensorimotor regions showed task-related seasonal patterns, with enhanced integration during longer daylight periods, potentially representing evolutionary adaptations optimizing motor performance. Abou Elseoud et al. [15] reported increased sensorimotor connectivity in SAD patients during winter, suggesting the clinical relevance of these seasonal changes. Athletic studies [14, 16, 18] revealed activity-dependent changes in sensorimotor connectivity, with Fitzgerald et al. [18] demonstrating that somatomotor networks were among the most affected by cumulative seasonal exposure in football athletes.
The SN, comprising the anterior insula and dorsal anterior cingulate cortex, demonstrates seasonal and temperature-related modulation. Granés et al. [10] investigated over 2000 children and found that higher ambient temperatures (17.7–25.5 °C) were associated with lower within-network connectivity in the SN, with the strongest effects observed the day before scanning. This suggests that thermoregulatory demands may compete with cognitive resources. Xu et al. [9] reported seasonal patterns in attention-related networks, though with different temporal profiles than the DMN and SMNs. Hu et al. [22] found that later scan times were associated with lower network efficiency and stability in dorsal attention and the SN, indicating circadian-seasonal interactions. Orban et al. [20] observed paradoxical reductions in global signal fluctuation from morning to evening, with widespread decreases in functional connectivity throughout the day affecting attention systems. Meyer et al. [6] demonstrated that attention-related activity peaked around the summer solstice, while working memory showed different seasonal timing, suggesting the functional specialization of AN seasonal responses. Abou Elseoud et al. [15] found heightened AN connectivity in SAD patients during winter, potentially indicating compensatory mechanisms during periods of reduced environmental stimulation.
VN connectivity shows seasonal variations associated with photoperiod changes. Abou Elseoud et al. [15] found increased connectivity in VN during winter in SAD patients compared to controls, interpreted as compensatory responses to reduced light input. Zhang et al. [11] demonstrated that VN connectivity was correlated with day length in longitudinal single-subject data. Orban et al. [20] reported that visual cortex connectivity decreased throughout the day, suggesting circadian modulation. Hu et al. [22] found that time of day, time of week, and time of year all affected VN properties, with weekend and summer scans showing distinct profiles.
Limbic network connectivity variations show functional significance for affective states. Racicot et al. [17] used 40–50 scanning sessions per participant to demonstrate that limbic network connectivity variations (involving amygdala and hippocampus) explained 15% and 11% of within-subject variance in positive and negative effects, respectively. Monroe et al. [19] identified increased central autonomic network (CAN) connectivity in soccer players exposed to repetitive seasonal impacts, particularly in left insula-orbitofrontal pathways linked to impaired cognitive control. Granés et al. [10] found temperature effects on hippocampal connectivity in children, indicating environmental sensitivity of limbic structures during development.
Across the included studies, certain consistent patterns emerge: (1) the DMN and SMNs show robust autumn peaks with summer minimums, closely tied to photoperiod [6, 9, 11]; (2) the SN demonstrates temperature sensitivity with inverse patterns to the DMN [10]; (3) the AN shows variable patterns that could reflect different functional roles [6, 20, 22]; (4) the VN responds to both seasonal photoperiod and daily circadian rhythms [11, 15, 20]; and (5) limbic networks show meaningful relationships with affective states across environmental fluctuations [17, 19]. These patterns indicate network-specific environmental sensitivities that may have adaptive and clinical implications.
Table 3 provides a systematic comparison of seasonal changes across different brain networks and their relationships with environmental variables by summarizing the key findings related to research objectives (1) and (2).
| Brain network | Peak season | Minimum season | Primary environmental driver | Correlation pattern | Studies (n/N) | Key regions affected/Functional implications |
| DMN | Autumn | Summer | Photoperiod duration | 12/16 | PCC, mPFC, angular gyrus/Memory, mood, self-referential processing | |
| SMN | Autumn | Summer | Photoperiod duration | 10/16 | M1, SMA, S1/Motor control, sensory processing | |
| SN | Summer | Winter | Ambient temperature | 6/16 | Anterior insula, dACC/Interoception, thermoregulation | |
| DAN | Winter | Summer | Inverse photoperiod | 4/16 | Superior frontal gyrus, supramarginal gyrus/Goal-directed attention | |
| VAN | Summer/Autumn | Winter | Mixed variables | 3/16 | Ventral frontal cortex, TPJ/Stimulus-driven attention | |
| VN | Summer | Winter | Photoperiod duration | 5/16 | V1, visual association areas/Visual processing, circadian regulation | |
| CAN | Post-season | Pre-season | Athletic exposure | No | 2/16 | PAG, hypothalamus, amygdala/Stress response, autonomic regulation |
Abbreviations: dACC, dorsal anterior cingulate
cortex; DAN, dorsal attention network; DMN, default mode network; M1, primary
motor cortex; mPFC, medial prefrontal cortex; PAG, periaqueductal gray; PCC,
posterior cingulate cortex; S1, primary somatosensory cortex; SMA, supplementary
motor area; SMN, sensorimotor network; SN, salience network; TPJ, temporoparietal
junction; V1, primary visual cortex; VAN, ventral attention network; VN, visual
network; CAN, central autonomic network.
Photoperiod emerged as an important environmental predictor of brain connectivity changes across the included studies. Xu et al. [9] reported that average day length was significantly associated with brain functional activities, with environmental factors, including day length, showing significant correlations with connectivity measures. Meyer et al. [6] demonstrated that photoperiod influences task-related brain activation patterns, with attention-related activity peaking around the summer solstice while working memory activation was highest during the autumn equinox. In dense longitudinal data, Zhang et al. [11] found that day length was associated with increased SMN and VN connectivity. While these studies establish photoperiod as a significant environmental factor, the specific nature of dose-response relationships and potential threshold effects require further systematic investigation.
Temperature effects were examined in two studies. Granés et al. [10] investigated ambient temperatures ranging from 17.7–25.5 °C during the week before scanning in 2229 children aged 9–12 years. Higher temperatures within this range were associated with lower within-network connectivity in the medial parietal network, salience network, and hippocampus, with the strongest effects observed the day before scanning (
Table 4 provides a comprehensive framework comparing different research approaches used in environmental neuroscience studies to systematically address research objective (3) regarding methodological strengths and limitations.
| Methodological aspect | Approach | Studies using | Advantages | Limitations | Recommendation level |
| Study design | Cross-sectional | 9/16 (56%) | Large sample sizes possible; Cost-effective; Broad population sampling; Seasonal comparisons across groups | Vulnerable to confounding; Cannot establish causality; Individual variability masking; Cohort effects | Moderate—Good for initial exploration |
| Longitudinal | 7/16 (44%) | Within-subject comparisons; Controls individual differences; Causal inference possible; Seasonal tracking | Expensive and time-consuming; Dropout concerns; Practice effects; Limited sample sizes | High—Preferred for causal inference | |
| Scanner technology | 3.0 Tesla | 15/16 (94%) | Optimal signal-to-noise ratio; Standardized protocols; Wide availability; Good connectivity detection | Susceptible to motion artifacts; Environmental interference; Cost considerations | High—Current standard |
| 1.5 Tesla | 1/16 (6%) | Lower cost; Reduced motion sensitivity; Historical data availability | Lower signal quality; Reduced connectivity sensitivity; Limited comparability | Low—Acceptable for specific contexts | |
| Sample size categories | Small (20–50) | 6/16 (38%) | Intensive phenotyping possible; Detailed individual analysis; Pilot feasibility; Dense sampling potential | Limited generalizability; Statistical power concerns; Effect size uncertainty; Outlier sensitivity | Moderate—Good for proof-of-concept |
| Medium (51–500) | 4/16 (25%) | Balanced power and feasibility; Subgroup analyses possible; Moderate generalizability | May miss small effects; Resource intensive; Recruitment challenges | High—Optimal balance | |
| Large ( |
6/16 (38%) | High statistical power; Excellent generalizability; Small effect detection; Population representation | Reduced individual detail; Quality control challenges; Expensive; Phenotyping limitations | High—Best for population inferences | |
| Environmental measurement | Direct individual measurement | 3/16 (19%) | Precise exposure assessment; Individual-specific data; Temporal accuracy; Dose-response relationships | Logistically complex; Expensive; Limited historical data; Participant burden | High—Most accurate approach |
| Regional/meteorological data | 13/16 (81%) | Historical availability; Cost-effective; Standardized measurements; Population-level inference | Individual exposure variation; Spatial resolution limits; Microenvironment differences; Timing mismatches | Moderate—Acceptable approximation | |
| Analysis approaches | Seed-based connectivity | 8/16 (50%) | Hypothesis-driven; Region-specific insights; Computational efficiency; Clinical interpretability | Limited to predefined regions; Potential bias; Network interactions missed; Seed placement sensitivity | Moderate—Good for targeted analysis |
| Independent Component Analysis | 6/16 (38%) | Data-driven approach; Whole-brain coverage; Network identification; Minimal assumptions | Component interpretation challenges; Reproducibility concerns; Statistical complexity; Parameter sensitivity | High—Comprehensive but complex | |
| Graph theory metrics | 4/16 (25%) | Network-level properties; Topological insights; Quantitative measures; Cross-study comparability | Abstract measures; Clinical translation difficulty; Threshold dependency; Computational complexity | Moderate—Advanced applications | |
| Control strategies | Basic demographics (age, sex) | 16/16 (100%) | Standard practice; Known confounders; Regulatory requirements | Insufficient for seasonal studies; Residual confounding; Interaction effects ignored | Minimum—Necessary but insufficient |
| Physiological controls (sleep, caffeine) | 4/16 (25%) | Seasonally-relevant factors; Mechanism identification; Confound reduction | Measurement challenges; Participant compliance; Cost increase; Temporal matching | High—Critical for interpretation | |
| Technical controls (phantom scans) | 2/16 (13%) | Scanner stability verification; Artifact identification; Quality assurance | Additional scanning time; Resource requirements; Limited availability | High—Essential for validity |
The methodological approaches employed across environmental neuroscience studies reveal some strengths and critical gaps that influence the interpretation and clinical translation of the results (Table 4). While most studies utilized appropriate 3.0T MRI systems and standard demographic controls, significant methodological heterogeneity limits direct cross-study comparisons. The predominance of regional meteorological data over individual environmental measurements introduces exposure misclassification that may attenuate true effect sizes. Most critically, only 13% of studies included technical controls such as phantom scans to verify scanner stability across seasons, despite evidence that environmental factors can influence scanner performance. The balanced representation of cross-sectional and longitudinal designs provides complementary perspectives, though the superior causal inference capabilities of longitudinal approaches make them preferable for establishing environmental-neural relationships. Sample sizes were appropriately distributed across small, medium, and large studies, though power calculations specific to seasonal effect sizes are rare. Future research should prioritize Tier 1 methodological requirements while incorporating Tier 2 enhancements to strengthen causal inference and clinical applicability.
Seasonal connectivity effects are comparable to those associated with psychiatric and neurological conditions [9], raising concerns for clinical neuroimaging if seasonal changes are not controlled. This could confound group comparisons or longitudinal assessments, potentially masking or artificially enhancing patient-control differences. These findings offer insights into mood disorder pathophysiology and intervention timing guidance. Understanding seasonal neural adaptations could inform educational practices, workplace policies, and clinical treatments, thereby optimizing year-round cognitive performance. The research reinforces the environmental neuroscience value for public health strategies, particularly regarding climate change’s impacts on the population’s cognitive function. Future research should standardize acquisition protocols, employ larger diverse samples, and control for biological and technical confounders to isolate environmental rhythm signatures and clinical implications.
This review synthesized findings from most available fMRI studies investigating how seasonal and circadian rhythms modulate brain functional connectivity. The evidence suggests that both macro-temporal (seasonal) and micro-temporal (circadian) environmental rhythms have measurable and sometimes profound influences on intrinsic brain network dynamics. These effects appear to be widespread across cognitive, sensory, and affective circuits, with implications for both healthy function and neuropsychiatric vulnerability. One of the most consistent findings was the seasonal modulation of the DMN, a system central to self-referential thinking and internal mentation [26]. Previous studies demonstrated that DMN connectivity fluctuates with seasonal shifts in photoperiod and solar exposure, reaching peaks in autumn and declining in summer [6, 9]. This observation aligns with prior research showing that mood, cognition, and the levels of neurochemicals such as serotonin vary seasonally in healthy individuals [27, 28]. Functional neuroimaging studies have linked reduced daylight exposure to diminished serotonin transporter binding [4], which may partially mediate seasonal fluctuations in mood and brain connectivity.
Circadian rhythms, intrinsic approximately 24-hour biological oscillations entrained by environmental cues, were also found to influence network architecture. Orban et al. [20] and Hu et al. [22] reported diurnal decreases in sensory and attentional network connectivity, demonstrating the effects of time of day on brain functional organization. These findings are consistent with studies in chronobiology showing circadian modulation of attention, executive function, and cortical excitability [29, 30]. Notably, Zhang et al. [11] and Racicot et al. [17] used high-density sampling to demonstrate that these effects are not merely interindividual but are observable within single participants across days and seasons.
Importantly, alterations in functional connectivity were also evident in non-clinical and developmental populations. Granés et al. [10] showed that higher ambient temperatures reduced connectivity in SN and medial parietal networks in children, supporting the notion that thermal stress may impair brain function in vulnerable populations. This complements emerging research in environmental neuroscience suggesting that heat exposure impairs working memory and executive performance via changes in neurovascular and metabolic efficiency [31]. Given that children’s brains are undergoing rapid development, seasonal and environmental variation may have long-lasting implications for educational and cognitive outcomes. The relationship between environmental modulation and psychiatric risk was particularly salient in studies of SAD. Abou Elseoud et al. [15] demonstrated enhanced connectivity in visual and attentional circuits during winter in SAD patients, which may represent compensatory mechanisms to reduce sensory input. Such alterations in connectivity may reflect a breakdown in the regulation of affective salience and cognitive control under seasonal stress. Moreover, clinical research has shown that seasonal fluctuations in brain activity may precede mood symptoms, suggesting a neurobiological diathesis that interacts with environmental context [4, 17].
In the domain of sports neuroscience, repeated subconcussive exposure across athletic seasons also altered intrinsic connectivity. Previous studies show DMN and AN changes in athletes over time [14, 18, 19]. These findings mirror neuropsychological evidence indicating that even subclinical head trauma can disrupt functional network efficiency and lead to cumulative neurocognitive deficits [32]. Given the seasonal structure of many athletic programs, it is crucial that future fMRI studies in sports consider time of year and accumulated exposure in their designs. However, these findings should be interpreted with caution. Di et al. [13] demonstrated that environmental variables such as weather conditions could predict imaging results using phantom and anatomical scans, suggesting that some seasonal fMRI patterns may result from scanner instability or technical drift. These artifacts can mimic neural activity if not rigorously controlled. This reinforces calls in the neuroimaging community for multi-site replication, routine phantom calibration, and the standardization of environmental variables during scanning [33].
In summary, the evidence underscores the dynamic interplay between environmental rhythms and brain functional architecture. While the precise mechanisms remain under investigation, likely contributors include photoperiod-induced hormonal shifts (e.g., melatonin, cortisol), seasonal changes in serotonin and dopamine function, and environmental influences on vascular tone and metabolism [34]. The findings have wide-reaching implications not only for clinical diagnostics and brain health but also for neuroimaging research protocols, which must increasingly account for these “invisible” environmental variables.
Despite growing evidence for environmental modulation of functional connectivity, the current literature has several limitations. First, there is considerable heterogeneity in methodology, including differences in sample size, scanner strength, preprocessing pipelines, and statistical thresholds. This limited our ability to make direct comparisons between studies and complicates meta-analytic synthesis. Second, although most studies controlled for major confounders such as age and gender, few of them addressed behavioral or physiological factors, such as sleep, caffeine intake, or mood at the time of scanning, which may themselves be seasonally modulated. Third, environmental data such as daylight or temperature were often inferred from regional averages and were not always verified at the individual level, introducing potential measurement bias. Fourth, the majority of studies used cross-sectional designs, which are prone to interindividual variability. While longitudinal studies offer greater sensitivity to change, only a subset of included studies adopted this design. Furthermore, dense-sampling studies, though informative, remain underpowered for generalization. Finally, very few studies included diverse age ranges, socioeconomic backgrounds, or non-Western populations, limiting the global applicability of the findings. There is also a notable gap in the integration of multimodal imaging or physiological metrics, such as melatonin levels or cortisol rhythms, which could clarify underlying mechanisms.
This review highlights that seasonal and circadian factors have a measurable influence on brain functional connectivity, as detected through rs-fMRI and task-based fMRI. The evidence spans across healthy and clinical populations, from children to adults, and includes both cross-sectional and longitudinal designs. Seasonal influences appear most prominently in the DMN, as well as the sensory and limbic networks, while circadian effects are evident in attention and sensorimotor systems. These findings underscore the need to account for time of year and time of day in basic and clinical neuroscience research. To advance this field, future studies should incorporate rigorous methodological controls, larger and more diverse cohorts, multimodal physiological measures, and standardized scan timing protocols. Doing so will help disentangle genuine neurobiological rhythms from technical artifacts and foster a more nuanced understanding of how the brain aligns with environmental cycles.
• Seasonal and circadian factors, particularly photoperiod and temperature, have clinically significant effects on brain functional connectivity across multiple networks, with magnitudes comparable to neurological and psychiatric disorders.
• The DMN and SMN show seasonal modulation with autumn peaks and summer minimums correlating with photoperiod, while salience networks demonstrate inverse temperature-related patterns.
• Time of day effects reveal paradoxical reductions in global signal and sensorimotor/visual connectivity from morning to evening, emphasizing the need to control scan timing in neuroimaging protocols.
• Scanner stability artifacts can mimic biological seasonal effects, necessitating rigorous technical controls, including phantom scans and multi-site replication, to distinguish genuine neurobiological rhythms from confounds.
• Seasonal connectivity variations have critical implications for clinical neuroimaging interpretation and research design, as uncontrolled seasonal factors may confound patient-control comparisons or mask treatment effects.
• Future research should prioritize longitudinal designs with dense sampling, individual-level environmental measurements, multimodal assessments, and standardized protocols accounting for seasonal and circadian timing.
This systematic review is based on previously published studies. The data used in this systematic review are available from the corresponding author upon reasonable request.
All elements of the study and subsequent write-up were carried out by the author [NAM]. The author read and approved the final manuscript. The author has participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
The author gratefully acknowledges the contributions of research assistants from the Diagnostic Radiography Technology (DRT) Department who supported the literature screening and data extraction stages of this review. Sincere appreciation is also extended to colleagues from the College of Nursing and Health Sciences for their valuable methodological guidance. The author also thanks Jazan University, Saudi Arabia, for providing the online resources and institutional support necessary to complete this research.
The author gratefully acknowledge the funding of the Deanship of Graduate Studies and Scientific Research, Jazan University, Saudi Arabia, through Project number: JU-202502140-DGSSR-RP-2025.
The author declares no conflict of interest.
Supplementary material associated with this article can be found, in the online version, at https://doi.org/10.31083/BJHM53980.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.