




 , Gaheon Lee 1, Kyung Hwa Kim 1,2,*
, Gaheon Lee 1, Kyung Hwa Kim 1,2,*
1 Department of Health Sciences, The Graduate School of Dong-A University, 49315 Busan, Republic of Korea
2 Department of Health Care and Science, College of Health Science, Dong-A University, 49315 Busan, Republic of Korea
Abstract
Mitochondria play crucial roles in maintaining health and influencing disease progression by acting as central regulators of cellular homeostasis and energy production. Dysfunctions in mitochondrial activity are increasingly recognized as key contributors to various pathologies, ultimately impacting healthspan and disease outcomes. However, traditional treatments often do not restore damaged mitochondria to a healthy state. Mitochondrial transplantation, a cellular organelle-based therapy in which mitochondria are introduced into a recipient, has emerged as a novel concept in next-generation therapeutics that overcomes the limitations of current cell-based treatments. This review highlights the unique properties of mitochondria as therapeutic agents, including their ability to restore cellular functions and treat a wide range of diseases. In this review, we focus on the unique role of mitochondria in the regulation of stem cell functions, including stem cell fate, self-renewal, and differentiation. Various perspectives have been explored to better understand mitochondrial transplantation therapy, which harnesses the capacity of mitochondria as living drugs in regenerative medicine, as an innovative strategy to bridge the gap between cell therapy and organelle-based treatments and overcome current clinical barriers.
Graphical Abstract
Keywords
- mitochondrial transplantation
- organelle transplantation
- mitochondrial dysfunction
- mesenchymal stem cell
- regenerative medicine
Mitochondria are unique organelles capable of self-renewal via biogenesis and elimination via degradation. Their dynamic turnover is vital for maintaining cellular health because it ensures the replacement of damaged or dysfunctional mitochondria [1, 2]. Disruption of this delicate balance can result in mitochondrial dysfunction which has far-reaching consequences for both cellular integrity and organismal health. Mitochondrial diseases arise primarily from mutations in either mitochondrial DNA (mtDNA) or nuclear DNA, with an estimated global prevalence of approximately 1 in 5000 individuals [3, 4, 5]. However, these disorders exhibit a wide spectrum of associated genetic mutations and clinical phenotypes, with considerable variability in disease onset and progression, making determination of an accurate prevalence rate challenging [6]. Furthermore, the heterogeneous presentation of symptoms among affected individuals complicates diagnosis, and as mitochondrial diseases are rare genetic disorders, large-scale research and clinical development remain limited [7]. Consequently, current therapeutic strategies for treating mitochondrial diseases focus primarily on managing symptoms and improving patient outcomes, as no definitive cure has been established to date.
Mitochondria are not static entities. Since the pioneering observations of mitochondrial dynamics in tissue culture by Lewis MR and Lewis WH [8] in 1914, advancements in imaging technologies have revealed the remarkable mobility and dynamic behavior of mitochondria within living cells [9, 10, 11]. Emerging evidence indicates that certain cells can export their mitochondria to neighboring cells, a phenomenon known as mitochondrial transfer [12]. Intercellular mitochondrial transfer occurs under physiological and pathological conditions [13]. Mitochondrial transfer has been observed through structures such as tunneling nanotubes (TNTs) [14], microvesicles (MVs) [15], and internalized free mitochondria [16, 17], enabling the restoration of damaged or dysfunctional cells by providing functional mitochondria.
As the diverse mechanisms of spontaneous mitochondrial transfer have been revealed, mitochondrial transplantation therapy has emerged as a revolutionary strategy in the field of regenerative medicine. Mitochondrial transplantation offers a promising solution to major challenges: the inability of traditional treatments to fully restore mitochondrial function and the persistent limitations of current cell-based therapies. Mitochondrial dysfunction is central to numerous diseases [18, 19]. However, conventional therapies often fail to repair or replace damaged mitochondria fully, resulting in an unmet need for targeted interventions. McCully’s research team first demonstrated the cardioprotective effect of mitochondrial transplantation [20], suggesting the potential application of this strategy in the treatment of various diseases.
This review highlights the potential of mitochondria as therapeutic agents, emphasizing the roles of mitochondria in restoring cellular function and treating a wide range of diseases. By focusing on their ability to regulate stem cell function and act as living drugs, we explore mitochondrial transplantation therapy as a promising strategy to bridge the gap between traditional cell-based therapies and next-generation organelle-based treatments.
Over the past few decades, regenerative medicine has remarkably advanced, offering transformative approaches for tissue repair and organ regeneration [21, 22]. Compared with traditional pharmaceuticals, cell-based therapies have distinct advantages, such as superior biocompatibility, enhanced bioavailability, and the ability to deliver greater therapeutic payloads [23]. Various cell-based treatments have demonstrated the potential to replace or repair damaged tissues and organs, showing great promise in preclinical and clinical settings [24].
Despite their transformative potential, cell-based therapies face significant factors that limit their widespread success. Some of these primary hurdles are low cell survival and poor functional integration after transplantation. For example, the damaged tissue microenvironments, which is characterized by inflammation and oxidative stress significantly impair the survival of mesenchymal stem cells (MSCs), as demonstrated by Kong et al. [25]. Immune rejection and the risk of graft-versus-host disease remain critical obstacles, particularly when allogeneic cells are used [26, 27]. Heterogeneity within stem cell populations further complicates therapeutic outcomes and often leads to inconsistent therapeutic efficacy. Concerns regarding tumorigenicity and uncontrolled differentiation, especially that of pluripotent stem cells, underscore the need for improved safety measures [23].
New approaches have recently been explored from a broader perspective to overcome these drawbacks and increase the accessibility of this technology to a broader population. Among these, organelle-based therapeutics have emerged as promising alternatives [28]. Notably, mitochondria are prominent among cellular organelles because of their remarkable ability to function semi-independently within the cell. Several unique characteristics distinguish mitochondria from other organelles, rendering mitochondria well suited as biopharmaceutical agents for therapeutic applications [29] (Fig. 1). Key advantages exist in the use of mitochondria in therapeutic contexts as organelle-based therapeutics to overcome some of the inherent limitations of traditional cell-based therapies.
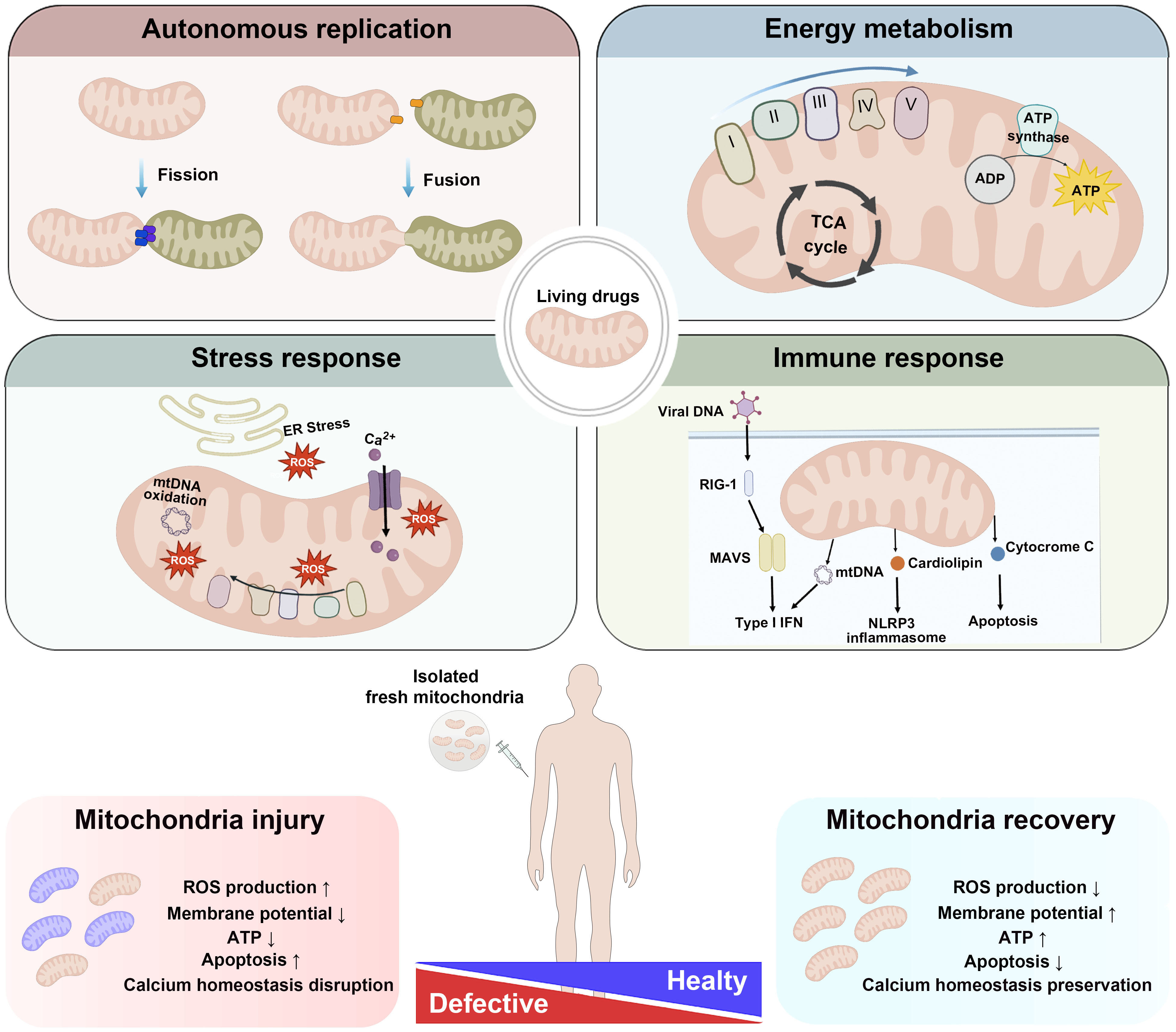 Fig. 1.
Fig. 1.
The unique properties of mitochondria as living drugs.
Mitochondria possess unique properties that make them promising candidates for
therapeutic applications in regenerative medicine. Unlike other organelles,
mitochondria contain their own genetic material and can undergo dynamic processes
such as division and fusion to adapt to cellular conditions and maintain
homeostasis. These organelles are also the primary sites of ATP production
through oxidative phosphorylation, supplying the energy necessary for cellular
functions. In addition to their role in energy metabolism, mitochondria play
critical roles in cellular stress responses by regulating ROS levels.
Additionally, they serve as key regulators of immune responses and are
intricately involved in the modulation of programmed cell death pathways. In
mitochondrial diseases, dysfunctional mitochondria fail to sustain normal
cellular processes, leading to pathological conditions. Healthy mitochondria that
are transplanted to a recipient have been shown to integrate into the host
mitochondrial network, thereby promoting tissue repair, enhancing cellular
survival, and facilitating regeneration. Given these multifaceted functions,
mitochondria themselves have emerged as potential “living drugs” in
regenerative medicine and precision therapy. This paradigm shifts underscores the
potential of mitochondria-based approaches to overcome the limitations of
conventional targeted therapies, paving the way for the next generation of
therapeutic strategies. Abbreviations: ER, endoplasmic reticulum; ROS, reactive
oxygen species; mtDNA, mitochondrial DNA; MAVS, mitochondrial antiviral signaling
protein; NLRP, NOD-like receptor protein; NLRP3, NOD-like receptor family, pyrin
domain containing 3; RIG-I, retinoic acid-inducible gene I. Arrows:
Mitochondria possess a unique intrinsic replication capacity, which enables them to proliferate independently of the host cell division cycle. This feature plays a crucial role in regulating mitochondrial dynamics, including fission, in which a single mitochondrion divides into multiple fragments, and fusion, in which individual mitochondria merges. This dynamic equilibrium is essential for cellular homeostasis. Mitochondrial fission facilitates the removal of damaged mitochondria, thereby contributing to mitochondrial quality control and overall cellular function [30]. The autonomous replication of mitochondria is facilitated by their own circular DNA (mtDNA) and dedicated replication machinery, enabling mitochondria to maintain their population and functionality even in nondividing or slowly dividing cells, such as neurons and muscle cells [31]. Mitochondrial fusion enables damaged mitochondria to merge with healthy mitochondria, facilitating functional restoration and enhancing overall mitochondrial efficiency [30]. This finely regulated fission–fusion mechanism plays a critical role in maintaining cellular homeostasis and has significant implications for long-term therapeutic strategies targeting mitochondrial dysfunction [32].
Mitochondria-based interventions may not require frequent replenishment because organelles can persist and function over extended periods. Mitochondria possess a remarkable ability to modulate their number through biogenesis, aligning with the metabolic demands of tissues. This adaptability is crucial for the maintenance of cellular energy homeostasis, particularly under various physiological and pathological conditions. In healthy tissues, mitochondrial biogenesis is tightly regulated to fulfill energy requirements [33, 34, 35]. For example, during endurance exercise, the mitochondrial content of skeletal muscle cells increases significantly, which enhances oxidative capacity and endurance. Regular aerobic exercise can reportedly lead to a 50–100% increase in mitochondrial density in muscle fibers, depending on the intensity and duration of training [36]. This augmentation leads to increased adenosine triphosphate (ATP) production, which is essential for sustained muscle contraction and performance.
Under pathological stress, such as stress due to ischemia or neurodegenerative
diseases, mitochondrial biogenesis may be impaired, leading to energy deficits
and cellular dysfunction [37, 38, 39, 40]. Therefore, therapeutic interventions that
stimulate mitochondrial biogenesis can counteract these deficiencies. For
example, in models of cardiac hypertrophy, increased mitochondrial biogenesis has
been associated with improved cardiac function and stress resistance [41].
Pharmacological agents such as Peroxisome proliferator-activated
receptor-
The extent of post-transplantation mitochondrial proliferation varies depending on factors such as the tissue type, extent of injury, and method of mitochondrial delivery. Although precise quantitative data are still being investigated, the therapeutic potential of mitochondrial transplantation is evident in various preclinical studies [42]. Ongoing research is aimed at optimizing transplantation techniques and understanding the long-term effects of the introduced mitochondria on host tissues.
In addition to their traditional role as powerhouses, mitochondria are recognized as central hubs of inter-organelle communication, that coordinate with various cellular compartments to maintain homeostasis and facilitate stress responses [43]. This communication occurs through direct physical interactions and signaling molecules, enabling dynamic responses to metabolic and environmental changes.
Recent studies have reported a key role of mitochondria in stress management in coordination with various organelles, including the nucleus [44], endoplasmic reticulum (ER) [45], and lysosomes [46]. Mitochondria-associated membranes serve as a physical platform for interactions between mitochondria and the ER. These physical contact sites facilitate signal transmission, thereby regulating a wide range of cellular processes associated with both organelles, including ER stress, mitochondrial dynamics, and apoptosis [47].
Historically, molecules released from stressed mitochondria have been considered solely proapoptotic triggers. Recent studies have revealed that these signals often initiate adaptive responses that promote cellular protection and resilience. This phenomenon is a hallmark of mitohormesis, a process in which mild mitochondrial stress induces signaling events [48]. Rather than being purely detrimental, signals released from damaged mitochondria often trigger protective cellular mechanisms that orchestrate complex and sophisticated adaptive responses. These signaling events, which are mediated by molecules such as reactive oxygen species (ROS), acetyl-CoA, mitochondria-derived peptides (MDPs), and mtDNA, can activate pathways that increase cellular resilience, promote repair, and restore homeostasis [44]. These signals from damaged mitochondria orchestrate complex protective mechanisms, emphasizing the sophisticated roles of these signals in cellular adaptation.
The unique ability of mitochondria to mediate cellular adaptation and protection underscores their potential in therapeutic strategies such as mitochondrial transplantation. By replacing diseased or aged mitochondria with healthy mitochondria, rejuvenating cellular functions and promoting regeneration at the whole-cell level is possible.
Mitochondria are unique organelles because of their circular double-stranded DNA (mtDNA), which is distinct from the nuclear DNA of the cell. This autonomous genetic material enables mitochondria to independently synthesize specific proteins essential for their function, including components of the electron transport chain and oxidative phosphorylation (OXPHOS). This autonomy is a key advantage when considering mitochondria as therapeutic agents in transplantation.
One of the most notable benefits of mitochondrial transplantation is its ability to bypass nuclear genomic alterations. Because mtDNA is independent of the nuclear genome, therapies targeting or replacing damaged mitochondria do not disrupt nuclear DNA, thereby avoiding the off-target effects or unintended genetic mutations often associated with nuclear genome editing technologies. Thus, mitochondrial transplantation is an inherently safe option for genetic therapy.
Additionally, autonomous mitochondrial DNA provides a direct avenue for treating diseases caused primarily by mutations or deletions in mtDNA, such as mitochondrial encephalomyopathy, lactic acidosis, and stroke-like episodes as well as Leber’s hereditary optic neuropathy [49]. By transplanting healthy mitochondria containing intact mtDNA, normal mitochondrial function can be restored and disease symptoms can be alleviated at the cellular level. In addition to inherited mitochondrial diseases, aging is strongly associated with mtDNA mutations and damage, which lead to mitochondrial dysfunction [50].
This capability is particularly advantageous because mitochondrial transplantation for aging and aging-associated diseases can be performed without immune suppression or complex genetic modifications. From targeted treatment of mitochondrial genetic disorders to more broad applications in regenerative medicine, the ability to repair or replace dysfunctional mitochondria without interfering with nuclear DNA represents a significant advancement in the development of safe and effective cellular therapies.
Mitochondria are central to cellular energy metabolism and supply the energy required for vital biological processes via ATP production. Given the vital role of mitochondria in energy production, it is not surprising that pioneering research on mitochondrial transplantation has been predominantly aimed at treating ischemic diseases in the context of cardiology and neurology to increase metabolic function. For example, in ischemic heart models, mitochondrial transplantation significantly increases intracellular ATP levels in cardiomyocytes and promotes tissue recovery [51]. Similarly, under neurodegenerative conditions, transplanted mitochondria show the potential to mitigate neuronal energy deficits and increase survival [52]. A comprehensive discussion of these therapeutic applications is also presented.
Ultimately, the critical role of mitochondria in energy metabolism extends beyond sustaining cell survival. If a sufficient number of transplanted mitochondria are adopted by the recipient cells, healthy mitochondria can facilitate seamless functional recovery in energy-deficient environments. Thus, targeting mitochondria to increase energy production offers a promising strategy for regenerating tissue, accelerating recovery, and improving the functionality of damaged tissues, with significant implications for regenerative medicine applications.
Owing to their small size and dynamic nature, mitochondria can traverse cellular compartments and deliver therapeutic agents, including proteins, RNA, DNA, and small molecules, directly into target cells. A critical aspect of mitochondrial transplantation is the ability to precisely target mitochondria to specific tissues or organs, thereby ensuring their bioavailability and maximizing their therapeutic efficacy. Given the role of mitochondria in cellular energy production, oxidative stress regulation, and signaling, the ability to accurately deliver mitochondria to damaged or energy-deficient tissues is fundamental for achieving clinical success.
Subsequent strategies have been devised to improve the targeting of transplanted mitochondria to living drugs. For example, chemically conjugating mitochondria with triphenylphosphonium results in the specific delivery of transplanted mitochondria into the brain meninges, but not into the brain itself [53]. Once mitochondria are transplanted, their bioavailability depends critically on their unique ability to functionally integrate into host cells, which distinguishes them from other cellular organelles. Unlike other organelles, mitochondria possess dynamic properties, such as fusion, fission, and biogenesis, enabling them to adapt and integrate seamlessly into the existing mitochondrial network of the host cell.
Mitochondrial biogenesis, the process of generating new mitochondria, further amplifies the therapeutic potential of mitochondria. Transplanted mitochondria can trigger biogenesis signals within host cells, increasing overall mitochondrial mass and improving cellular resilience under stress [54]. The mechanisms underlying this process are discussed in detail in subsequent sections.
This remarkable ability to functionally integrate with host cells underscores the uniqueness of mitochondria as therapeutic tools. Unlike other organelles, which play static roles or require external support, mitochondria dynamically engage in cellular processes, adapt to the host environment, and provide sustained therapeutic benefits [55]. This dynamic integration highlights the role of mitochondria as not only cellular powerhouses but also active participants in cellular repair and regeneration.
Clinical trials exploring mitochondrial transplantation therapy are advancing rapidly, with companies such as Cellvie, Mitrix Bio, and Minovia Therapeutics making efforts to commercialize allogeneic and autologous mitochondrial products. Prebanked allogeneic mitochondria sourced from healthy donors undergo rigorous isolation, purification, and expansion to ensure a sufficient quantity of viable mitochondria for therapeutic use.
Cellvie is at the forefront of the development of mitochondrial transplantation therapies, which aim to restore cellular energy metabolism and improve tissue function through mitochondrial transplantation. Notably, this company is actively investigating therapeutic strategies for ischemia-reperfusion injury, a condition commonly associated with stroke and organ transplantation [20, 56]. Mitrix Bio is advancing bioreactor-based technology for large-scale mitochondrial cultivation. By utilizing cell lines with high energy-producing capacity, mitochondria proliferate in a controlled bioreactor environment, after which they are isolated and purified, producing components termed “mitlets”. These mitlets are currently being explored as a potential therapeutic approach for various diseases, including Alzheimer’s disease and immunosenescence. Pelletier et al. [57] reported that extracellular vesicles (EVs) containing functional mitochondria derived from platelets can enhance mitochondrial function and support metabolic activity in recipient cells. Additionally, Minovia Therapeutics specializes in autologous mitochondrial transplantation. For mitochondrial augmentation therapy, mitochondria are extracted from a patient’s own cells, expanded ex vivo, and subsequently reinfused into the body. This personalized therapeutic approach is particularly advantageous for rare mitochondrial disorders such as Pearson syndrome, as this minimizes the risk of immune rejection. Recently, Minovia Therapeutics conducted a study in which mitochondria from a healthy donor were transplanted into patients with mtDNA defects. These mitochondria were introduced into the patient’s hematopoietic stem cells, leading to increased energy production and improved cellular function [58, 59].
Similar to MSC therapy, in mitochondrial transplantation therapy, mitochondria are prepared for allogeneic or autologous transplantation, in which they are administered to unrelated recipients to restore cellular bioenergetics and function. Although the exact mechanisms of mitochondrial transplantation are not yet fully understood, clinical studies are underway to verify the therapeutic efficacy of mitochondrial transplantation in various diseases (Table 1, Ref. [60, 61, 62, 63, 64, 65, 66]). In particular, mitochondrial transplantation has been proposed as an innovative strategy to restore cellular energy metabolism and enhance the functional recovery of damaged tissues. Ongoing clinical trials primarily utilize autologous mitochondria for direct injection into the lesion site, and large-scale clinical protocols are still lacking. Since most mitochondrial diseases are studied in the form of case studies, large-scale controlled studies should be conducted to establish protocols and determine effective treatment strategies for each disease. Therefore, it is necessary to establish a thorough design system for future clinical trials. For example, an optimization process to evaluate the appropriate mitochondrial donor site, injection route, dose, and number of treatments in each disease should be performed. Solving the above problems can overcome the limitations of existing cell-based therapies and present a new paradigm for regenerative medicine.
| Disease | Source | Donor | Administration | Phase | Status | Enrollment | Experimental design | Efficacy evaluation indicators | NCT number |
| Cerebral Ischemia | Autologous | Muscle tissue adjacent to the surgical access site | Brain artery via microcatheter during reperfusion | 1 | Recruiting | 20 | Exp; Autologous mitochondria Transplantation | Cerebral angiography, CT scan, Peripheral blood study, Muscle biopsy, Brain MRI | NCT04998357 (Walker M et al. [60], 2024) |
| Myocardial Infarction, Myocardial Ischemia, Myocardial Stunning | Mitochondria; Pectoralis muscleExosome; Mesenchymal stem cells | Intracoronary and Intra-myocardial | 1/2 | Unknown | 20 | Exp; Exosome therapy/Mitochondria therapy/Co-transplantation of mitochondria and exosome therapyPC; Placebo | Creatine kinase (CK), Cardiac troponin T, On pump duration, ECMO duration, SPECT, NYHA, CMR | NCT05669144 (Tafti A [61], 2022) | |
| Extracorporeal Membrane Oxygenation Complication | Skeletal muscle of the chest wall | Intracoronary | N/A | Recruiting | 16 | Exp; Autologous mitochondria Transplantation | Ejection fraction, Ability to be separated from ECMO support | NCT02851758 (Emani S [62], 2023) | |
| Repetition Failure | Bone mesenchymal stem cells | Direct injection in the oocytes | N/A | Unknown | 50 | Exp; Autologous mitochondria transplantationAC; Intracytoplasmic sperm injection | Live birth rate, Clinical pregnancy rate, Number of oocytes retrieved, Fertility rate, Normal fertility rate, Good quality embryo rate | NCT03639506 (Deng Y [63], 2018) | |
| Infertility | Ovarian stem cells | Direct injection in the oocytes | N/A | Completed | 59 | Exp; ICSI with mitochondriaAC; ICSI without mitochondria ICSI | Rate of ongoing pregnancy, Morphological, Morphokinetic criteria and in Preimplantation Genetic Screening | NCT02586298 (Labarta E et al. [64], 2017) | |
| Low Prognosis After Repeated IVF Failure | Urine derived stem cells | Direct injection in the oocytes | N/A | Enrolling by invitation | 40 | Exp; MT transfer | Live birth rate | NCT06020742 (Han H [65], 2023) | |
| Polymyositis, Dermatomyositis | Allogeneic | Umbilical cord-derived mesenchymal stem cells (PN-101) | Intravenous | 1/2 | Completed | 9 | Exp; Low/Intermediate/High dose group | International Myositis And Clinical Studies group-Total Improvement Score (IMACS-TIS), Response rate of IMACS-TIS, Changes of Core Set Activity Measures (CSAM), Changes of Cutaneous Dermatomyositis Disease Area and Severity Index (CDASI), Changes of Peak Pruritus Numeral Rating Scale (PPNRS) | NCT04976140 (Paean Biotechnology Inc [66], 2024) |
ICSI, Intracytoplasmic sperm injection; Exp, Experimental; AC, Active comparator; PC, Placebo comparator; IVF, in vitro fertilization; N/A, not applicable; MT, mitochondria; MRI, magnetic resonance imaging; ECMO, extracorporeal membrane oxygenation; SPECT, Single Photon Emission Computed Tomography; NYHA, New York Heart Association Functional Classification; CMR, cardiovascular magnetic resonance.
Although the precise mechanisms governing mitochondrial transplantation are not fully understood, recent studies have identified key pathways through which transplanted mitochondria exert their effects (Fig. 2).
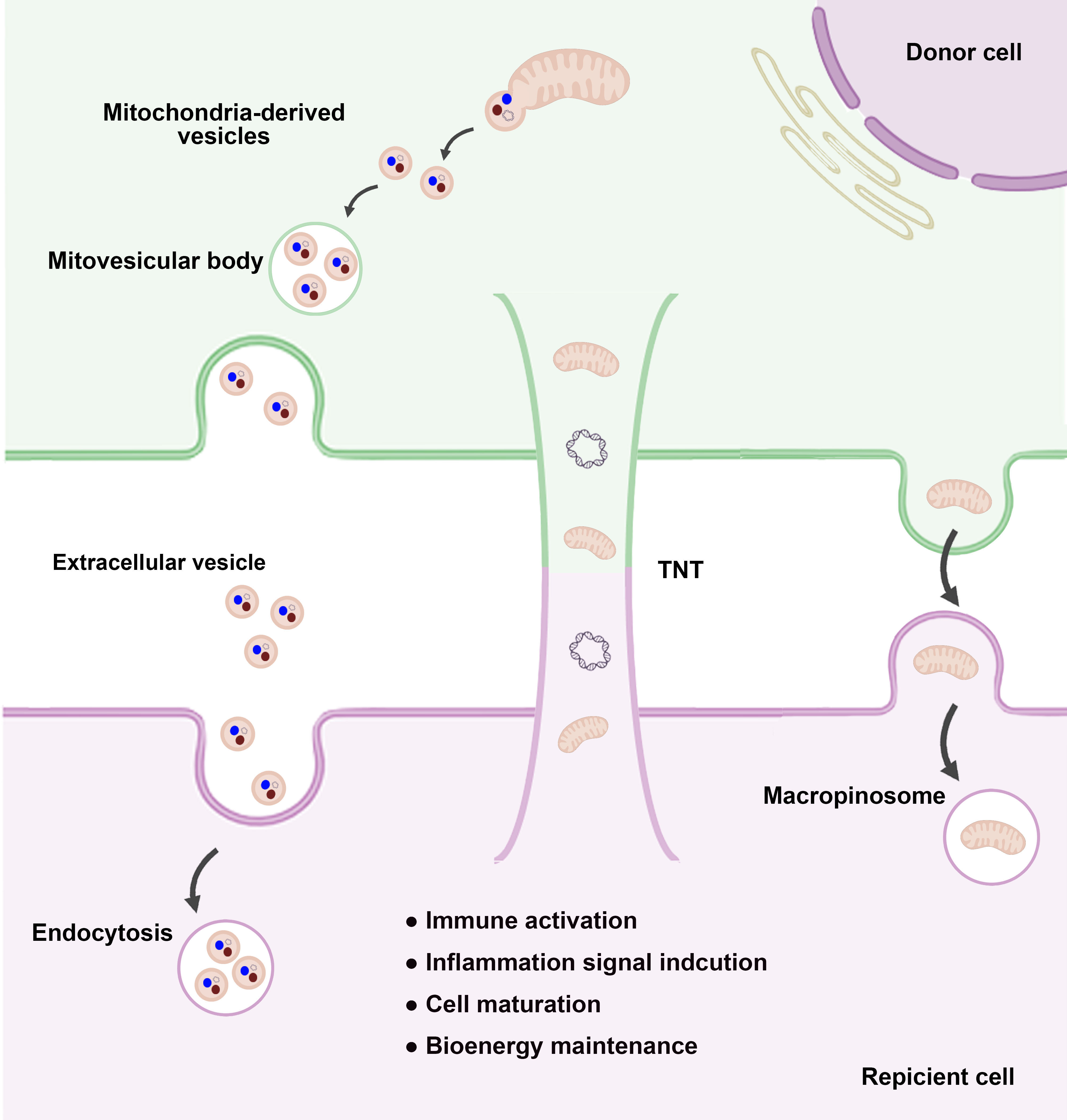 Fig. 2.
Fig. 2.
Mitochondrial transfer pathways and biological effects. Mitochondria can be transferred between cells through various mechanisms and play crucial roles in intercellular communication and cellular homeostasis. One such mechanism involves MDVs, which contain either whole mitochondria or mitochondrial components. These vesicles are secreted into the extracellular space and incorporated into EVs, which are subsequently internalized by recipient cells via endocytosis. Another major pathway is TNT-mediated transfer, in which mitochondria are directly transferred through filamentous structures that physically connect donor and recipient cells. This mechanism enables the exchange of functional mitochondria, supporting cellular repair and metabolic recovery in damaged recipient cells. Additionally, micropinocytosis-mediated transfer involves the uptake of free mitochondria by recipient cells through macropinosome formation, facilitating mitochondrial acquisition and cellular adaptation. These diverse mitochondrial transfer mechanisms have been shown to modulate immune responses, regulate inflammatory signaling, and contribute to cell maturation and bioenergetic homeostasis. Understanding these processes provides valuable insights into mitochondrial dynamics and their therapeutic potential in regenerative medicine and disease treatment. Abbreviations: MDVs, mitochondrial-derived vesicles; EVs, extracellular vesicles; TNTs, tunneling nanotubes.
Mitochondria are highly dynamic organelles that respond to cellular damage and exhibit remarkable adaptability. Recent studies confirmed that mitochondria can be transferred between cells under both physiological and pathological conditions. Intercellular mitochondrial transfer was first observed in 2004, when mammalian cells were shown to exchange mitochondria via TNTs [14]. These thin tubular structures, composed of the plasma membrane and actin filaments, connect cells over long distances, facilitating the horizontal transfer of organelles, such as mitochondria, as well as proteins, DNA, RNA, and lipids. Similarly, osteocytes reportedly transfer mitochondria to metabolically stressed cells through dendrite-like membranous protrusions, resulting in restored metabolic function [67].
Intercellular mitochondrial transfer is tightly regulated by interactions between the ER and mitochondria. Other routes of mitochondria transfer include the use of extracellular MVs [13, 68], which encapsulate whole mitochondria and deliver them to target cells.
In addition to strategies that utilize their membranous structure, mitochondria can be exchanged extracellularly in two steps: extrusion and internalization. Under conditions of mitochondrial stress, cells extrude dysfunctional mitochondria via a process called mitoptosis [69] or release intact mitochondria as danger signals [70]. Subsequently, neighboring cells engulf these mitochondria, triggering proinflammatory signaling and promoting cell maturation.
The internalization of free mitochondria, a process referred to as mitochondrial
transformation, was first reported in 1982 [71]. In 2006, Spees et al.
[72] discovered that A549
Nonetheless, recent evidence suggests that free mitochondria can be taken up through micropinocytosis [16, 17], in which large vesicles called macropinosomes engulf extracellular fluid and organelles. Studies have suggested that regulatory factors such as syncytins [76] and the integrity of the mitochondrial outer membrane [17] play critical roles in this process.
Advanced strategies, including surface modification with cell-penetrating peptides (e.g., Pep-1 and trans-activator of transcription (TAT) peptide) [77, 78], and the use of biocompatible carriers, or photothermal nanoblades [79, 80], have been developed to increase mitochondrial internalization. These methods are crucial for expanding mitochondrial transplantation for diverse clinical applications, such as cardiac ischemia and neurodegenerative diseases, where mitochondria restore bioenergetics and repair damaged tissues [75].
Standardizing mitochondrial transfer protocols and enhancing delivery efficiency are critical for understanding the full therapeutic potential of mitochondrial transplantation. The emerging field of biomaterial–mitochondrial research has led to significant advancements, particularly in strategies aimed at improving mitochondrial internalization. These strategies include (1) surface modification of mitochondria with cell-penetrating peptides or cationic coatings [79]; (2) encapsulation of mitochondria within biocompatible carriers to protect and enhance delivery [81]; (3) pressure-driven [82] or magnet-driven [83] techniques to precisely direct mitochondrial localization; and (4) physical principle-based methods, such as photothermal nanoblades, which transiently permeabilize cell membranes to facilitate mitochondrial uptake [79, 80].
Recent studies have highlighted mitochondria as secretory organelles capable of releasing a wide range of bioactive molecules that profoundly influence cellular signaling and function [44]. These secreted factors, including proteins, mtDNA, and metabolites, act as paracrine signaling molecules that modulate cellular processes under both normal and stressful conditions. Although research on paracrine activation by transplanted mitochondria is still in its early stages, several mitochondria-derived components have been implicated in this process.
During mitochondrial transplantation, or in response to cellular stress, transplanted or damaged mitochondria may release mtDNA into the cytosol or extracellular space (Fig. 3). The released mtDNA interacts with pattern recognition receptors, such as Toll-like receptor 9 (TLR9) and the cyclic GMP-AMP synthase-stimulator of interferon genes (cGAS-STING) pathway, triggering downstream signaling cascades [84]. In a pioneering study, Islam et al. [68] observed human mtDNA in lipopolysaccharide-injected mice following the administration of human bone marrow-derived MSCs, providing direct evidence for mtDNA transfer via EVs.
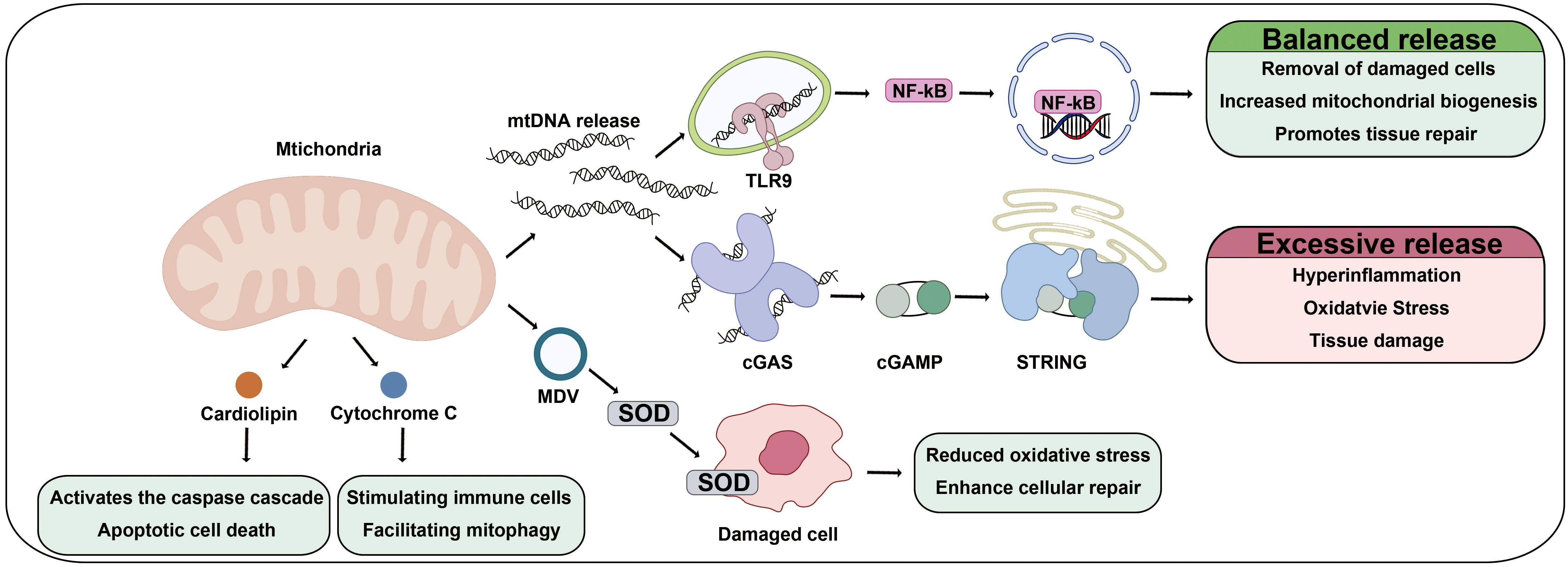 Fig. 3.
Fig. 3.
Therapeutic mechanisms of mitochondria under pathological conditions. Under pathological conditions, components derived from transplanted mitochondria exert therapeutic effects by modulating immune responses, reducing oxidative stress, and promoting tissue regeneration. Mitochondria-released mtDNA is recognized by TLR9 on immune cells, initiating signaling pathways that regulate inflammation and facilitate tissue repair. Additionally, mtDNA binds cGAS, leading to the production of cyclic GMP-AMP (cGAMP) and subsequent activation of the STING pathway, which enhances immune responses and supports cell survival. Furthermore, transplanted mitochondria release MDVs containing SOD, which alleviates oxidative stress by neutralizing ROS in recipient cells. MDVs also deliver mitochondrial proteins and lipids, contributing to mitochondrial biogenesis and metabolic restoration. The components released from transplanted mitochondria serve as bioactive mediators that promote cellular regeneration, metabolic adaptation, and immune modulation, ultimately supporting tissue repair and maintaining cellular homeostasis in damaged tissues. Abbreviations: TLR9, Toll-like receptor 9; cGAS, cyclic GMP-AMP synthase; STING, stimulator of interferon genes; SOD, superoxide dismutase.
Intriguingly, the immune response initiated by mtDNA release involves a delicate balance between beneficial and potentially harmful effects [85]. On the positive side, mtDNA-triggered signaling facilitates the clearance of damaged cells, enhances mitochondrial biogenesis, and supports tissue repair by promoting controlled inflammation. Moderate activation of the TLR9 and cGAS-STING pathways resolves tissue damage and promotes the functional integration of transplanted mitochondria into host cells. However, excessive mtDNA release or prolonged activation of these pathways can lead to hyperinflammation, oxidative stress, and tissue damage [86]. Furthermore, damage-associated molecular patterns (DAMPs), including cardiolipin and cytochrome C released from compromised cells, and mtDNA, activate the innate immune system and modulate damage repair mechanisms. Cardiolipin, a phospholipid located in the inner mitochondrial membrane, plays a crucial role in immune activation by stimulating immune cells to initiate inflammatory responses and by facilitating mitophagy for the clearance of dysfunctional mitochondria [87]. Additionally, during cellular damage, an increase in mitochondrial membrane permeability leads to the release of cytochrome C, which subsequently activates the caspase cascade, triggering apoptotic cell death [88]. In summary, mitochondrial damage-associated molecules not only elicit immune and inflammatory responses but also play pivotal roles in orchestrating tissue damage repair processes.
When healthy mitochondria are transplanted, these risks are mitigated by the intrinsic properties of functional mitochondria, which help regulate oxidative stress and minimize excessive immune activation. This controlled inflammatory response creates an environment conducive to tissue repair and regeneration, highlighting the therapeutic advantages of healthy mitochondria for transplantation.
Another prominent paracrine mechanism involves the release of mitochondrial-derived vesicles (MDVs). MDVs are lipid-encased structures containing mitochondrial proteins, lipids, and other bioactive molecules capable of modulating signaling pathways in recipient cells. For example, MDVs reportedly deliver antioxidant enzymes such as superoxide dismutase (SOD) to nearby damaged cells, thereby reducing oxidative stress and enhancing cellular recovery [16].
As described above, mitochondrial signaling molecules initiate and modulate multiple signaling pathways for homeostasis. When healthy MSCs are cocultured with recipient cells, ROS from recipient cells can stimulate MSCs, increasing mitochondrial transfer to recipient cells through TNTs [89]. In contrast, pretreatment of MSCs with ROS scavengers such as N-acetyl-L-cysteine inhibits mitochondrial transfer and biogenesis. Collectively, this evidence underscores the role of mitochondrial signaling molecules as potent mediators of paracrine signaling during mitochondrial transplantation.
The paracrine effects of transplanted mitochondria represent a powerful mechanism for promoting tissue repair and immune modulation. Key components, such as mtDNA, MDVs, and ROS, coordinate to facilitate intercellular communication, enhance mitochondrial dynamics, and support tissue regeneration. However, further mechanistic studies are needed to elucidate the precise roles of and interactions between these factors during mitochondrial transplantation. The development of strategies to optimize these paracrine effects is critical for advancing the clinical application of this promising therapeutic approach.
Recently, the pivotal role of mitochondria in the regulation of stem cell function has gained significant attention [90]. Mitochondria play multiple roles in stem cell maintenance, differentiation, and therapeutic applications. This mitochondria-linked signaling is increasingly understood to be central to stem cell biology, offering new insights into the roles of mitochondrial signaling in both physiological and therapeutic contexts.
Given the crucial impact of mitochondria on stem cell fate, the notion that mitochondria act as a crucial link between stem cell therapy and organelle-based treatments is compelling. Stem cells are an advantageous source of healthy functional mitochondria for therapeutic applications. Although current studies on interventions to enhance mitochondrial health are limited, the application and development of such technologies in stem cell culture hold considerable promise. By integrating these approaches, stem cells can be optimized as an ideal source of high-quality and functionally superior mitochondria for therapeutic applications.
Mitochondrial metabolism is a critical determinant of stem cell fate and maintenance, and distinct metabolic profiles are observed at different stages of development and differentiation (Fig. 4A). Stem cells at early developmental stages, including MSCs [91], embryonic stem cells [92], and induced pluripotent stem cells [93], rely predominantly on glycolysis rather than OXPHOS.
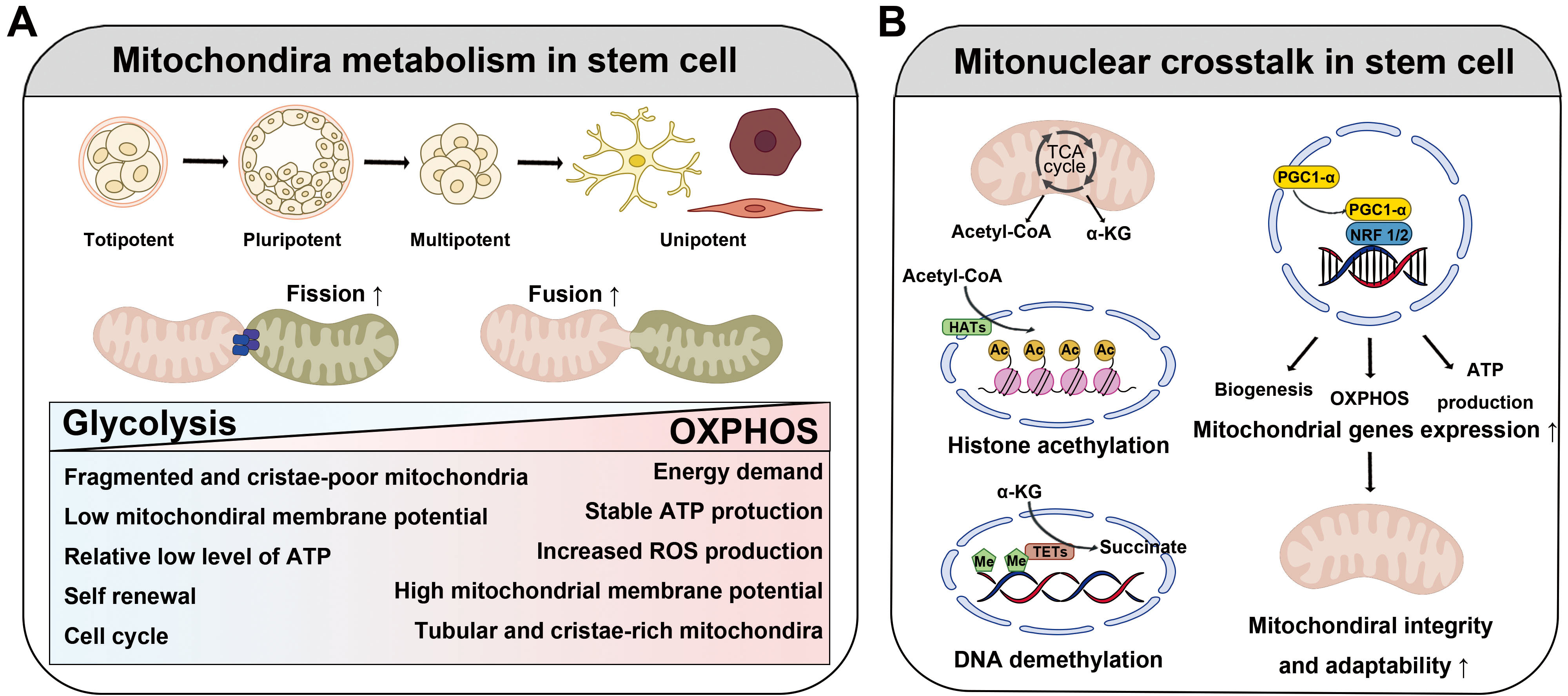 Fig. 4.
Fig. 4.
The function and role of mitochondria in stem cell fate
determination. (A) Mitochondria are key regulators of the development and
differentiation of stem cells. In their undifferentiated stage, mitochondria rely
mainly on glycolysis, and they are essential for maintaining the pluripotency of
stem cells by rapidly supplying energy even in hypoxic environments. When stem
cell differentiation begins, metabolic activity switches to OXPHOS. During this
period, mitochondria mature rapidly and actively induce fusion and division. This
process requires high energy, and the activation of OXPHOS contributes
significantly to cell fate determination. (B) Mitochondria communicate closely
with the nuclear genome and are involved in stem cell regulation. Mitochondrial
metabolites affect histone deacetylation and DNA demethylation, and nuclear
respiratory factors such as NRF1 and NRF2 also regulate mitochondrial function
and maintain stem cell characteristics. The regulation of mitochondrial nuclei is
closely linked to stem cell function, and understanding these interactions can
improve the efficiency of mitochondria-based treatments. Abbreviations: OXPHOS,
oxidative phosphorylation; NRF1/2, nuclear respiratory factors 1 and 2;
PGC-1
Although no definitive explanation for why stem cells prefer glycolysis exists, several possibilities have been proposed. One explanation for this finding is that stem cells often reside in hypoxic environments, such as bone marrow, where oxygen levels are low. Under such conditions, compared to OXPHOS, glycolysis is a more suitable energy-generating pathway that does not depend on oxygen availability [94]. Another possibility is that glycolysis generates significantly fewer ROS than does OXPHOS. By minimizing ROS production, glycolysis protects stem cells from oxidative damage and ensures genomic stability and long-term viability [90, 95]. In addition to energy production, glycolysis provides essential anabolic intermediates such as nucleotides, amino acids, and lipids. These intermediates are crucial for supporting the anabolic pathways necessary for stem cell self-renewal and progeny generation (see Suda et al. [95] and the references therein). Accordingly, glycolysis serves as a key process for biosynthesis, which is vital for maintaining stem cell populations and ensuring their proliferative capacity.
As stem cells differentiate, they undergo a metabolic shift toward OXPHOS [96], with elongated mitochondria exhibiting increased crista density. For example, MSC-derived osteoblasts and adipocytes rely heavily on OXPHOS to fulfill the energy demands of bone formation and lipid metabolism [97, 98]. Conversely, chondrocytes retain a glycolytic phenotype that favors the anabolic pathways necessary for cartilage maintenance. These dynamic shifts in mitochondrial metabolism underscore the intricate interplay between mitochondrial function and stem cell fate. Understanding the mechanisms governing these transitions is critical for optimizing stem cell-based therapies.
Coordination between the mitochondrial and nuclear genomes, known as mitonuclear
crosstalk, plays a fundamental role in stem cell regulation (Fig. 4B). This
communication ensures that mitochondrial function aligns with cellular metabolic
and transcriptional demands, thereby influencing the fate and behavior of stem
cells. Although intriguing, the mechanism by which mitochondria communicate with
the nucleus to maintain homeostasis remains unclear. However, some evidence
suggests that mitochondrial metabolites may modulate epigenetic regulation of
stem cell function [90]. For example, mitochondrial metabolites, such as
Another possibility is that stem cells are regulated by nuclear transcription factors. Stem cells require dynamic balance between glycolysis and OXPHOS to fulfill their metabolic demands during proliferation and differentiation. Nuclear respiratory factors 1 and 2, respectively (NRF1 and NRF2) play critical roles as key regulators of mitochondrial functions [100]. These transcription factors drive the expression of genes associated with mitochondrial biogenesis, OXPHOS, and ATP production, ensuring that mitochondrial activity is tightly coupled to the cellular metabolic state. Notably, NRF1- and NRF2-mediated transcription not only supports energy production but also reinforces mitochondrial integrity and adaptability, which are essential for stem cell maintenance and lineage specification. This regulatory axis highlights the intricate interplay between nuclear signaling pathways and mitochondrial function in stem cell biology.
Increased levels of PGC-1
Mitochondrial dysfunction is a critical factor that limits the regenerative
potential of stem cells, particularly with repeated passaging or aging.
Addressing this challenge requires innovative approaches to preserve and enhance
mitochondrial functionality. Restoring mitonuclear signaling through targeted
interventions, such as activating PGC-1
One potential approach involves leveraging MDPs such as humanin [105, 106] and mitochondrial open reading frame of the 12S rRNA type-c (MOTS-c) [107, 108]. These peptides play protective and regulatory roles in mitochondrial homeostasis and cellular metabolism. They may increase mitochondrial biogenesis, reduce oxidative stress, and modulate metabolic pathways, all of which are critical for maintaining stem cell function.
Another promising candidate is resveratrol, a polyphenolic compound known to
activate PGC-1
Although direct evidence of these approaches in stem cells remains limited, studies in other systems suggest that such approaches may hold promise in counteracting the mitochondrial dysfunction associated with aging or long-term culture when applied to stem cell culture systems. These interventions could pave the way for optimizing stem cell-based therapies by preserving the regenerative potential and enhancing the therapeutic efficacy of stem cells. Further research is needed to elucidate the mechanisms by which these strategies influence mitochondrial function and stem cell biology and ultimately improve outcomes in regenerative medicine.
Over the past few decades, mitochondrial transplantation therapy has garnered considerable attention. Several companies and research institutions have pioneered efforts to commercialize both allogeneic and autologous mitochondrial products. These advancements have led to the development of prebanked allogeneic mitochondria sourced from healthy donors via isolation, purification, and culture to generate sufficient quantities of viable mitochondria for therapeutic use. Despite these significant challenges, several critical issues must be addressed to increase the safety, efficacy, and scalability of mitochondrial transplantation therapies.
With advancements in single-mitochondrion sequencing technologies, it has become evident that mitochondria, even those within a single cell, exhibit diverse genetic and functional properties [111]. This heterogeneity includes variations in mtDNA mutations, heteroplasmy levels, and respiratory efficiency. These findings underscore the critical importance of selecting mitochondria with superior bioenergetic capacity and minimal mutational burden for therapeutic applications [111].
These insights into mitochondrial diversity highlight the need for careful consideration of variations during transplantation. For example, selecting populations with minimal mtDNA mutations is crucial to ensure the efficacy and safety of transplantation because mitochondria with fewer mutations are more likely to retain optimal respiratory function and bioenergetic capacity. In addition, identifying and isolating mitochondria tailored to the specific metabolic demands of the recipient tissue could significantly enhance therapeutic outcomes. In this context, mitochondria suitable for high-energy tissues, such as cardiac or neural tissues, may differ functionally from those required for less metabolically active environments. Furthermore, the age of the donor tissue can be a critical factor because mitochondria from younger donors generally exhibit greater respiratory efficiency, reduced oxidative stress, and fewer mtDNA mutations than those from older donors. These factors are essential for optimizing the success of mitochondrial transplantation therapy.
Notably, mitochondria are immune-privileged. Ramirez-Barbieri et al. [112] reported that neither syngeneic nor allogeneic mitochondria trigger significant acute, chronic, direct, or indirect immune responses. In this context, mitochondrial transplants appear to be immunologically well tolerated, regardless of their genetic origin. Moreover, in a study involving rats, dramatic improvements in survival and enhanced neurological recovery were observed even when allogeneic mitochondria were transplanted immediately after cardiac arrest [113]. These results suggest that mitochondrial transplantation, even transplantation of mitochondria from a genetically different donor, can sufficiently restore mitochondrial function to prevent extensive tissue damage and promote recovery in acute conditions such as cardiac arrest.
Mitochondrial function can be compromised at various stages of life, particularly in developmental and age-related contexts [114]. Congenital mitochondrial disorders arise from genetic mutations in mtDNA or nuclear DNA, leading to abnormal mitochondrial function and disrupted development. As individuals age, mitochondria naturally accumulate damage, resulting in reduced efficiency, increased susceptibility to oxidative stress, and increased risk of disease [114]. These findings highlight the potential of allogeneic mitochondrial treatment as a therapeutic option for severe mitochondrial dysfunction.
However, the following findings have driven the continued exploration of autologous mitochondria in regenerative medicine: (1) allograft rejection has been observed following a single injection of allogeneic mitochondria owing to the presence of foreign mtDNA or mitochondrial proteins, leading to immune responses and rejection [115] and (2) autologous mitochondrial transplants prevent the need for haplotype matching or familial donors, simplifying the transplantation process and minimizing the ethical concerns associated with donor sourcing and genetic inheritance [116, 117]. Notably, mitochondrial transplantation is distinct from mitochondrial replacement therapy, which involves transferring mitochondria or genetic material from a donor to an individual, raising questions about genetic identity and classification of the procedure. Despite these distinctions, both approaches offer valuable insights into potential treatments for mitochondrial dysfunction; however, ethical and immunological challenges remain.
In cases where mitochondria are not severely compromised, autologous mitochondrial transplantation is generally preferred over allogenic mitochondrial transplantation because of its immune compatibility and ethical clarity. In cases of severe mitochondrial dysfunction or congenital disorders, allogeneic transplantation may offer a more effective solution, although immune rejection remains a potential risk factor. In addition, recent gene editing techniques have been successful in editing mtDNA [118, 119]. These techniques have shown successful mitochondrial gene editing and may contribute to future therapeutic strategies by targeting genes that cause mitochondrial immunogenicity. Further advancements in immune modulation techniques are essential to improve the viability of allogeneic mitochondrial transplantation for broader clinical applications.
Despite extensive efforts to characterize functional mitochondria fully, their properties remain dependent on the cell culture conditions or the conditions of the tissue from which they are derived. A fundamental question arises regarding whether mitochondria function in the same manner as they do in vivo, or whether culture conditions influence their bioactivity. To date, several potential sources of mitochondria for transplantation, each with distinct advantages and limitations, have been explored.
Mitochondria present distinct energy demands and metabolic characteristics that are inherently influenced by the tissue of origin [120]. For example, mitochondria derived from metabolically active tissues, such as the heart and skeletal muscle, demonstrate high efficiency in ATP production, making them particularly suitable for therapeutic applications requiring robust energy supply. Conversely, mitochondria isolated from neural tissue are specialized for neuroprotection and intracellular signaling, whereas those derived from hepatocytes play crucial roles in detoxification and lipid metabolism. Consequently, the selection of an optimal mitochondrial source necessitates a comprehensive evaluation of the metabolic requirements and functional demands of the recipient tissue to ensure therapeutic efficacy and compatibility.
Since the first demonstration of mitochondrial transfer from MSCs to mitochondria-depleted cells, which restored aerobic respiration [72], MSCs have been regarded as one of the most promising renewable sources of exogenous mitochondria [68, 121]. As described above, compared with mitochondria derived from other cells, MSC-derived mitochondria possess several key advantages, including high metabolic activity, low immunogenicity, and regenerative potential. In particular, because MSCs can be isolated from diverse adult tissues, they also reduce ethical concerns and immunogenic consequences.
However, the functional properties of MSCs are highly variable, and depend on their culture and isolation. For example, heterogeneity within MSC populations poses challenges because MSCs comprise diverse subtypes, including pericytes and adventitial cells, each with distinct biological roles. Pericytes are more closely associated with neoangiogenesis, whereas adventitial cells contribute to bone formation [122, 123]. Additionally, the culture conditions used for ex vivo expansion may affect the functionality of MSCs. As MSCs age in vitro, they become flattened and enlarged, indicating cellular senescence, which typically begins at passage five [124]. Moreover, prolonged culture reduces proliferation and differentiation potential, leading to diminished bioenergetic efficiency, lower ATP production, and increased oxidative stress [125]. Maintaining stemness and bioenergetic efficiency over multiple passages is essential for ensuring the quality of MSC-derived mitochondria.
Considering these factors, the selection of an optimal mitochondrial source for clinical transplantation should be accompanied by a rigorous evaluation of therapeutic efficacy. For example, mitochondria derived from cardiac or skeletal muscle tissues, characterized by their high ATP production capacity, may be particularly suitable for the treatment of cardiovascular disorders. In contrast, mitochondria isolated from neural tissue, with their specialized role in mitigating oxidative stress and promoting neuroprotection may offer greater therapeutic potential for neurodegenerative diseases, as these mitochondria play specialized roles in mitigating oxidative stress and promoting neuroprotection. Future research should focus on systematically characterizing mitochondrial sources and developing standardized transplantation protocols to ensure reproducible and consistent therapeutic outcomes across diverse patient populations.
Other potential mitochondrial sources have also been investigated. A previous study has demonstrated the therapeutic effects of the transplantation of mitochondria from highly metabolic tissues, such as skeletal muscle [126]. Notably, recent study has investigated the impact of blood-derived mitochondria collected using minimally invasive methods [127].
To optimize mitochondria for therapeutic applications, several key areas, such as the standardization of MSC culture and preconditioning strategies through pharmacological agents or genetic interventions, require further research. Hence, defining a characterization strategy for identifying heterogeneous mitochondria with superior functions for transplantation is essential. Understanding the mechanistic underpinnings of mitochondrial therapies is essential to unlock the full therapeutic potential of mitochondria and ensure the efficacy and consistency of mitochondria as independently functioning organelles capable of driving cellular repair and regeneration.
Mitochondrial transplantation has emerged as a groundbreaking therapeutic
approach to address tissue damage and metabolic disorders. Among mitochondria
derived from various sources, mitochondria derived from MSCs have garnered
significant attention owing to their unique properties. These include intrinsic
anti-inflammatory capabilities, which suppress inflammation at the
transplantation site, and low immunogenicity, which reduces the risk of immune
rejection, thereby improving therapeutic outcomes. Recent studies have
demonstrated the efficacy of mitochondrial transplantation in various disease
models. For example, intravenous administration of mitochondria isolated from
MSCs exhibited therapeutic effects in an acute respiratory distress syndrome
model, in which it significantly reduced histopathological damage, inflammation,
and the levels of cytokines such as tumor necrosis factor-
Despite this therapeutic potential, mitochondrial transplantation faces
significant challenges related to mitochondrial internalization and functional
integration. The efficiency with which transplanted mitochondria are taken up by
recipient cells varies greatly depending on the cell type, and transplanted
mitochondria must integrate seamlessly into the existing mitochondrial network to
participate in metabolic and bioenergetic functions [131]. To overcome these
challenges, technologies for mitochondrial transfer and integration have been
developed. Technologies that enhance membrane receptor interactions through
nanoparticles and electroporation-mediated techniques have been reported to
improve mitochondrial entry into recipient cells [117]. In addition, genetic
engineering strategies that induce overexpression of mitochondrial biogenesis
regulators such as PGC-1
Although these findings highlight the promise of mitochondrial transplantation, further research is required to optimize delivery methods, enhance mitochondrial survival and integration, and minimize immune responses to identify its full potential as a highly effective and personalized therapeutic strategy in regenerative medicine.
Mitochondrial transplantation has emerged as a promising therapeutic approach in regenerative medicine. However, its clinical application remains limited due to several challenges, including low engraftment efficiency, quality control issues, and ethical concerns. The engraftment rate of transplanted mitochondria plays a pivotal role in determining therapeutic efficacy. However, studies have reported that in practice, the engraftment efficiency is relatively low, with a significant proportion of transplanted mitochondria being rapidly degraded or eliminated [75, 133]. This low engraftment rate not only diminishes therapeutic potential but also hampers the integration of transplanted mitochondria into the recipient’s endogenous mitochondrial network. Therefore, optimizing methodologies to enhance the stability and retention of transplanted mitochondria in vivo is imperative. In addition to engraftment challenges, mitochondrial quality control represents another critical factor. Mitochondria isolated from cells are prone to rapid functional deterioration, including loss of membrane potential, reduced ATP production, and increased oxidative stress, all of which can compromise their therapeutic efficacy and viability upon transplantation [134]. Recent studies have increasingly emphasized the significance of mitochondrial quality control (MQC) systems in maintaining cellular homeostasis [135]. Elucidating the key regulatory factors involved in MQC is expected to play a pivotal role in the development of novel therapeutic strategies targeting mitochondrial dysfunction. Thus, the systematizing of mitochondrial quality assessment is essential to ensure the consistency and reliability of transplanted mitochondria. The establishment of rigorous quality control criteria prior to transplantation is necessary to minimize variability and enhance clinical efficacy.
Furthermore, mitochondrial transplantation raises significant ethical concerns owing to the absence of standardized regulatory frameworks. The long-term stability and therapeutic outcomes of this approach remain inadequately characterized, indicating potential risks of unforeseen adverse effects [136]. In particular, if transplanted mitochondria can exhibit maternal inheritance, their impact on subsequent generations warrants ethical scrutiny. Therefore, comprehensive long-term monitoring is essential to evaluate safety and efficacy before widespread clinical implementation. Moreover, to address ethical considerations, it is crucial to establish fair access to treatment, secure informed consent from mitochondrial donors, and mitigate the risks associated with commercialization. Despite ongoing ethical and regulatory challenges, the limitations of mitochondrial transplantation can be addressed through the development of internationally standardized guidelines and legal frameworks. In addition to these scientific, ethical, and regulatory challenges, the economic feasibility of mitochondrial transplantation must also be considered to ensure successful clinical translation. Key cost drivers include mitochondrial isolation and purification, delivery technologies, and compliance with regulatory requirements, all of which demand significant investment. While mitochondrial transplantation may offer long-term cost benefits compared to existing cell-based therapies, the initial treatment cost and accessibility remain challenges. Therefore, development of scalable and automated manufacturing processes, along with policy-level support such as public health insurance coverage and R&D funding, will be essential for broader clinical adoption. Addressing these aspects will help establish mitochondrial transplantation as a viable and sustainable strategy in regenerative medicine. The establishment of a global database to monitor clinical applications and prevent unethical practices will further contribute to ensuring the responsible advancement of mitochondrial transplantation therapy.
Recent advances in mitochondrial biology have fundamentally transformed our understanding of these organelles, shifting a purely energy-centric perspective to a perspective that recognizes mitochondria as the central hubs of intracellular and intercellular signaling, a concept now referred to as mitochondria-centric signaling. This paradigm shift underscores the notion that the roles of mitochondria extend far beyond their role as cellular powerhouses; mitochondria are dynamic regulators of cellular homeostasis, health, and disease.
Mitochondria-dependent signaling pathways, which are active under both physiological and pathological conditions, influence a wide range of cellular processes, including calcium homeostasis, immune responses, and cellular stress adaptation. These pathways govern critical cellular decisions and have profound implications for organismal health, positioning mitochondrial dysfunction as a central contributor to various diseases such as neurodegenerative disorders, cardiovascular diseases, and cancer. Moreover, the ability of mitochondria to communicate with the nucleus and other cellular compartments highlights the integral roles of mitochondria in maintaining cellular homeostasis and responding to environmental stressors. Mitochondria-related signaling pathways are essential for cellular adaptation and survival under stress, providing promising therapeutic targets for disease management and intervention.
The restoration of mitochondrial function through strategies, such as mitochondrial transplantation, enhanced biogenesis, and improved quality control mechanisms, has demonstrated the potential to rejuvenate cellular function and advance regenerative medicine. These approaches offer novel therapeutic possibilities for addressing aging- and mitochondria-related diseases, replacing or repairing damaged cells, and restoring tissue function.
As our understanding of mitochondrial signaling dynamics continues to deepen, mitochondrial transplantation has emerged as a promising therapeutic frontier. By leveraging the unique properties of mitochondria, innovative strategies can be developed to repair and rejuvenate damaged tissues, offering transformative solutions for some of the most challenging diseases at present.
HRS and GHL contributed to the creation of figures and tables and participated in drafting and revising the manuscript. KHK, the corresponding author, contributed to the conception and structure of the review, participated in drafting and revising the manuscript, and supervised the overall work. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
Not applicable.
Not applicable.
This work was supported by a grant from the National Research Foundation of Korea (RS-2023-00243300) and by Dong-A University, both awarded to KHK.
The authors declare no conflict of interest.
During the preparation of this work the authors used ChatGPT 3.5 in order to check spell and grammar. After using this tool, the authors reviewed and edited the content as needed and took full responsibility for the content of the publication.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.