









1 Department of Dermatology, Second Affiliated Hospital, Zhejiang University School of Medicine, 310009 Hangzhou, Zhejiang, China
2 Department of Plastic Surgery, Second Affiliated Hospital, Zhejiang University School of Medicine, 310009 Hangzhou, Zhejiang, China
†These authors contributed equally.
Academic Editor: Ranieri Bizzarri
Abstract
Background: Artemis belongs to the SNM1 gene family, and plays a role in repairing ionizing-radiation-induced DNA double-strand breaks and variable (diversity) joining recombination. S534, S538, S516, S645 represent four most rapid phosphorylation sites in Artemis, and serine phosphorylation at amino acid 516 is closely associated with activation. Artemis mutation is perceived as contributing to Omenn syndrome, which manifest features of severe combined immunodeficiency disease, associated with lymphadenopathy, hepatosplenomegaly, erythroderma and baldness. In addition, Artemis phosphorylated at serine 516 (Artemis S516-P) was expressed in scalp hair follicles (HF) as well as other skin appendages, and its expression level is important to mouse hair cycling. However, whether Artemis participated in the regulation of HF growth still unclear. Methods: Using immunofluorescence double-staining, we assessed the association between Artemis S516-P with proliferation, apoptosis, and differentiation markers in normal adult anagen scalp HF. Results: The results of double-staining immunofluorescence revealed overlapping expression pattern for Artemis S516-P and keratin16, similar pattern for c-myc and p21, while presenting opposite trends for keratin 10, phospho-p53, Bax, Bcl-2 and keratin 14. Conclusions: Our study provides the clues that Artemis may play roles in regulation of differentiation, proliferation, apoptosis and cell cycling during HF growth and development.
Keywords
- phospho-Artemis (Serine516)
- hair follicle
- immunofluorescence
- double-staining
The Artemis gene was first identified in patients with radiation-sensitive
severe combined immunodeficiency disease (RS-SCID). Artemis mutations or
deficiency in RS-SCID patients appear to be associated with abnormal variable
(diversity) joining (VDJ) recombination, leading to arrested lymphocyte
maturation, consequent immune deficiencies and high sensitivity to ionizing
radiation (IR) [1, 2]. Artemis belong to the SNM1 gene family, which
possesses a conserved DNA-binding
Artemis is dually regulated by DNA-PKcs and the ATM protein kinase. ATM phosphorylation of Artemis is involved in protein ubiquitination, cell cycle regulation, maintenance of telomere function, and chromatin restructuring during various cellular stress responses [12, 13, 14, 15]. Both Artemis and ATM constitute the slow repair pathway for DSB repair. Artemis, regulated by both ATM and DNA-PKcs, plays a role in G1 phase of cell cycle via non-homologous end joining (NHEJ) pathways, while Artemis regulated by ATM alone acts on G2 phase through the homologous recombination pathway [16]. The repair of DSBs occurs mainly in highly dense heterochromatin areas, such as the telomere [15, 17]. Artemis can also promote p53 degradation by E3 ubiquitin ligase, and participate in apoptosis regulation and cell cycle progression by inhibiting p53 phosphorylation through DNA-PKcs [18, 19]. In addition to repairing DNA damage directly, Artemis may also participate in regulating cell cycle checkpoint and signal transduction pathways.
Artemis mutation is involved in the pathogenesis of Omenn syndrome [20, 21], which is characterized by SCID and baldness. Our previous studies showed that Artemis phosphorylated at serine 516 (Artemis S516-P) was detected in the inner and outer hair root sheaths of growing hair follicles (HF), hair shaft progenitor cells, and also the hair matrix (germ cells) near the papilla in human scalp skin [22], and the expression level of Artemis varies during mouse hair cycling [23]. HF germ cells are responsible for early changes in HF growth independent of HF bulge activity. In anagen HF, various cells comprising the inner root sheath and hair shaft proliferate and differentiate from the HF germ cells, while the outer hair root sheath differentiates from the HF bulge cells [24, 25]. Schuler et al. [26] showed that the accumulation of DNA damage-induced chromatin alterations, ultimately determined the biological outcome of HF stem cells. Therefore, Artemis may be involved in the early signaling of transform telogen to anagen in HF. In fact, Artemis S516-P was located mainly close to the epidermis in club hair, germ cells and cap, a region of abundant stem cells, during telogen in epilation-induced hair cycling in mice. Therefore, Artemis S516-P may contribute to HF development, though the underlying mechanisms remain unclear. Interestingly, our previous study showed that Artemis S516-P was expressed mainly in the cytoplasm in HF cells, while weakly expressed in the nucleus of hair bulb cells and hair outer root sheath basal cells [22], indicating its role as a signaling molecule in regulating HF cell proliferation and differentiation. To clarify these mechanisms, we investigated the co-expression of Artemis S516-P with various markers of proliferation, apoptosis, and differentiation in normal adult scalp.
Normal scalp tissue was obtained from seven Chinese subjects (mean
Anti phospho-p53, Bcl-2, Bax, keratin 10, keratin 14, keratin 16, p21, and c-myc antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Goat anti-rat-rhodamine and goat anti-rabbit-fluorescein isothiocyanate (FITC) antibodies were purchased from Dako Cytomation (Glostrup, Denmark). 4’,6-diamidino-2-phenylindole, (DAPI) was purchased from Sigma-aldrich (St Louis, MO, USA). Rabbit anti-Artemis S516-P polyclonal antibodies were prepared by Hangzhou HuaAn Biotechnology Company (Hangzhou, China). The sequence of the synthetic Artemis phosphorylation at serine 516 was: VAGGS (p) QSPKLFS (p denotes the phosphorylation site). Antibody specificity was confirmed in Hela cells after transfection with the Artemis mutation fragment (D37N-413), a competitive inhibitor serine 516 phosphorylation of Artemis [27].
Immunofluorescence double-staining for Artemis S516-P and markers of
proliferation, differentiation and apoptosis was carried out in human anagen HF
as follows. Normal scalp
specimens embedded in OCT were serial-sectioned at 5–8
The Artemis (serine 516) antibody was diluted 1:500, antibodies to phospho-p53, Bcl-2, Bax, keratin 10, keratin 14, keratin 16, p21, and c-myc were diluted 1:200. Each sample was stained at least 3–4 times to ensure consistent results.
Phospho-p53 (serine 15) expression was detected in the outer hair root sheath, the inner root sheath, and the cortex of the middle HF. Artemis S516-P expression was strongly positive in the middle and outer parts of the outer hair root sheath. These two proteins showed opposite expression patterns in the same locations; high Artemis S516-P expression co-localized with weak phospho-p53 (serine 15) expression, and vice versa (Fig. 1). In hair bulb, Phospho-p53 (serine 15) staining was more intensive, but staining for Artemis S516-P became less intensive. Negative controls were presented in Supplementary Fig. 1A.
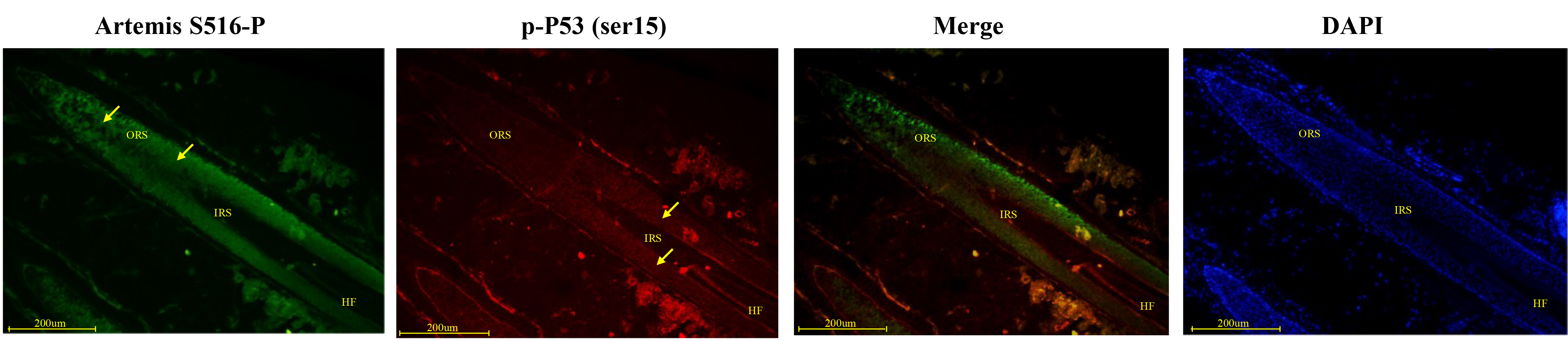 Fig. 1.
Fig. 1.Immunofluorescence double-staining for Artemis S516-P and phospho-p53 in human growing scalp HF. In situ immunofluorescence double-staining of cryostat sections of scalp from healthy donors. Phospho-p53 staining was observed in the outer root sheath lining, inner root sheath and cortex of the middle HF. Artemis S516-P staining was strongly positive in the middle and outer parts of the outer root sheath. Strong Artemis S516-P expression was associated with weak phospho-p53 expression, and vice versa. Artemis S516-P stained green; DAPI nuclear staining was blue; phospho-p53 stained red; merged staining was yellow. Positive staining was highlighted by arrows. Abbreviations: ORS, outer root sheath; IRS, inner root sheath; HF, hair follicle.
Artemis S516-P was strongly expressed in the middle and upper segments of HF, and its expression gradually reduced closer to the hair bulb. However, Bax expression increased as Artemis S516-P expression decreased (Fig. 2). Negative controls were presented in Supplementary Fig. 1A.
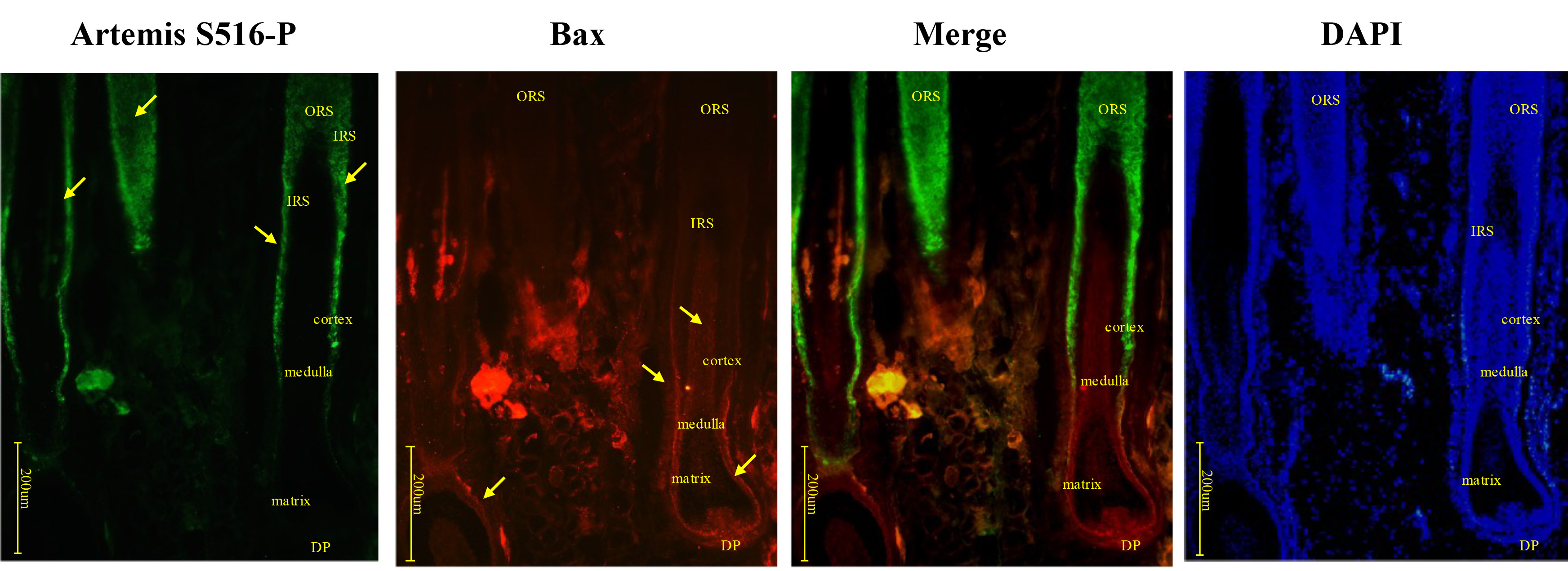 Fig. 2.
Fig. 2.Immunofluorescence double-staining for Artemis S516-P and Bax in human growing scalp HF. In situ immunofluorescence double-staining of cryostat sections of scalp from healthy donors. Artemis S516-P staining was strongly positive in the middle and upper segments of the HF, and gradually decreased near the hair bulb. Bax staining gradually increased closer to the hair bulb. Artemis S516-P stained green; DAPI nuclear staining was blue; Bax stained red; merged staining was yellow. Positive staining was highlighted by arrows. Abbreviations: ORS, outer root sheath; IRS, inner root sheath; DP, dermal papilla.
The expression patterns of Artemis S516-P and Bcl-2 resembled those of Artemis S516-P and Bax; Artemis S516-P expression gradually weakened nearer the hair bulb, while Bcl-2 expression gradually increased (Fig. 3). Negative controls were presented in Supplementary Fig. 1A.
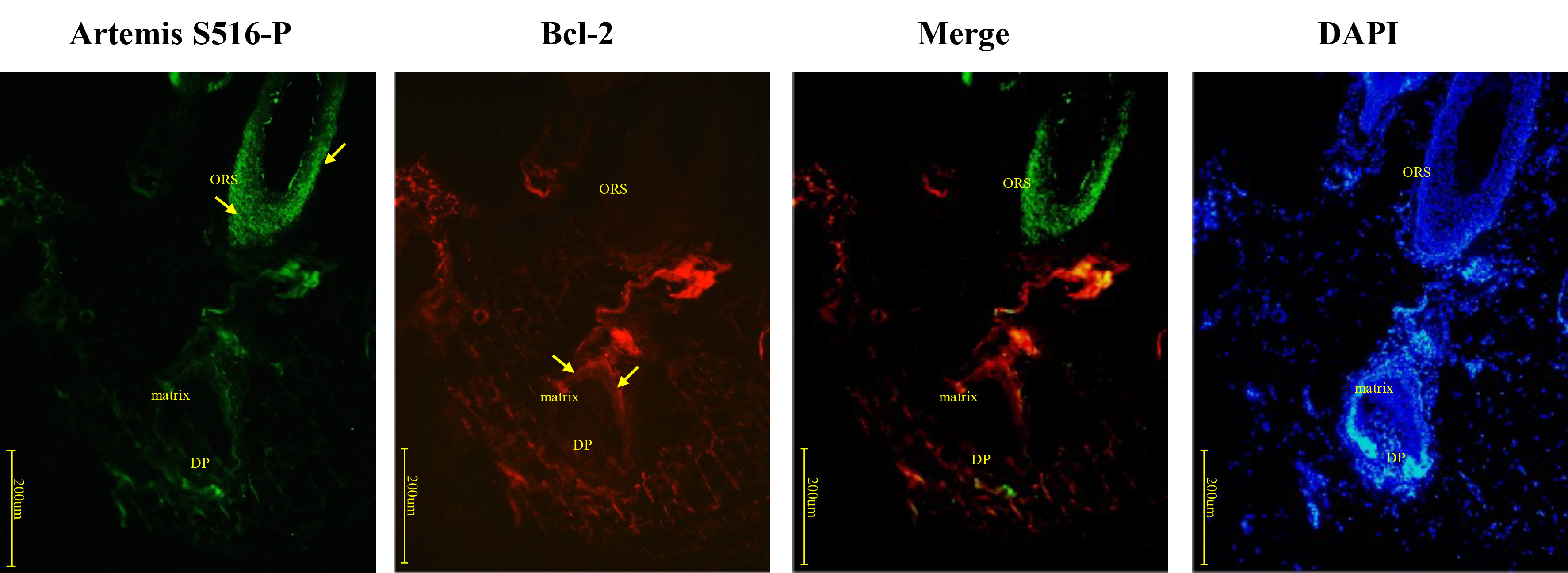 Fig. 3.
Fig. 3.Immunofluorescence double-staining for Artemis S516-P and Bcl-2 in human growing scalp HF. In situ immunofluorescence double-staining of cryostat sections of scalp from healthy donors. Artemis S516-P stained strongly positive in the middle and upper segments of the HF, and gradually decreased near the hair bulb, while Bcl-2 expression gradually increased. Artemis S516-P stained green; DAPI nuclear staining was blue; Bcl-2 stained red; merged staining was yellow. Positive staining was highlighted by arrows. Abbreviations: ORS, outer root sheath; DP, dermal papilla.
Artemis S516-P and keratin 10 presented contrasting expression patterns in the upper and middle sections of the HF. Artemis S516-P was expressed mainly in cells inside the HF, especially in outer hair root sheath cells in the middle and upper parts of the follicles, and was expressed only weakly in Malpighian layer. Keratin 10, however, was expressed mainly in Malpighian cells located in the epidermis between HF, sebaceous glands and sweat glands (Fig. 4). In addition, we also found that Artemis S516-P was widely expressed in dermal components, which was consistent with Artemis’ function. Since Artemis acts as a multifunctional protein involved in regulating cell cycle checkpoints, chromatin remodeling and cellular stress response. Therefore, normal dermal fibroblasts show Artemis expression. Negative controls were presented in Supplementary Fig. 1B.
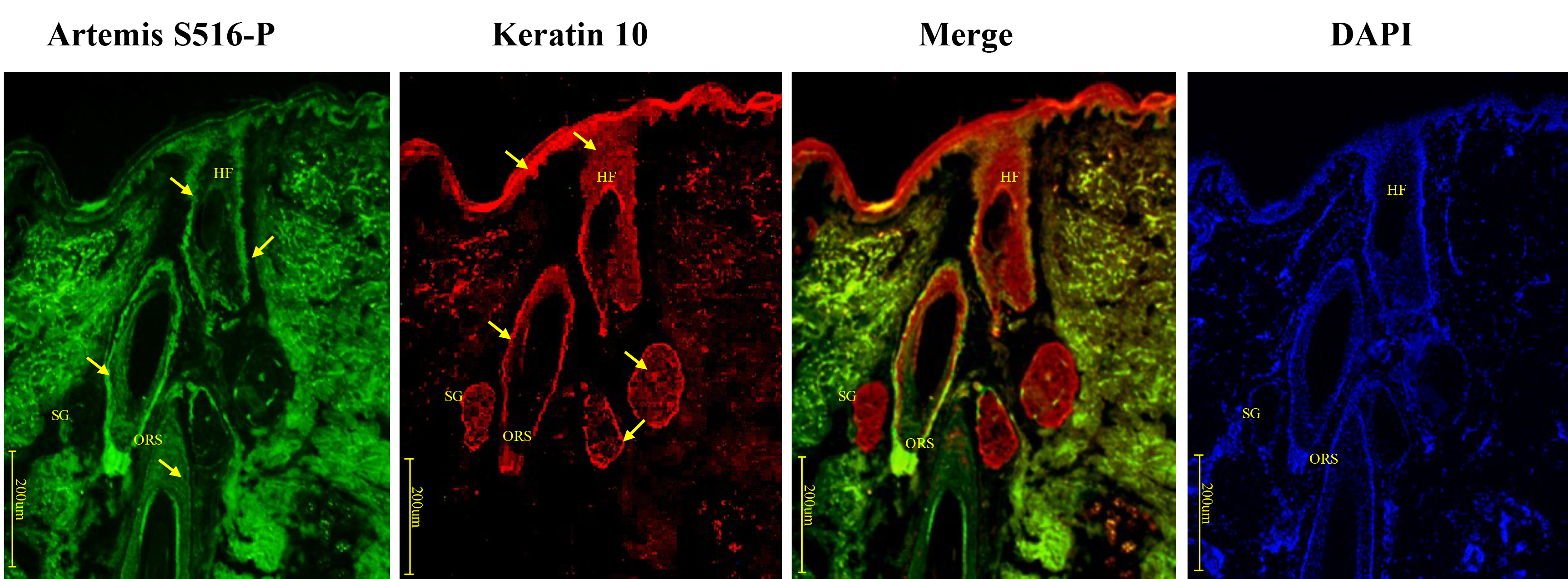 Fig. 4.
Fig. 4.Immunofluorescence double-staining for Artemis S516-P and keratin 10 in human growing scalp HF. In situ immunofluorescence double-staining of cryostat sections of scalp from healthy donors. Artemis (serine 516) and keratin 10 showed opposite staining patterns in the upper and middle sections of the HF. Artemis S516-P showed strong staining along the outer hair root sheath cells in the middle and upper parts of the HF, and weaker staining in other keratinocytes, while keratin 10 stained strongly in keratinocytes beside the HF, such as the epidermis between HF, sebaceous glands, and sweat glands. Artemis S516-P stained green; DAPI nuclear staining was blue; keratin 10 stained red; merged staining was yellow. Positive staining was highlighted by arrows. Abbreviations: ORS, outer root sheath; HF, hair follicle; SG, sweat gland.
Artemis S516-P and keratin 16 expression patterns showed close overlap. Both showed strong positive expression in the middle and upper sections of the HF outer root sheath, and weak expression in the hair matrix, medulla, cortex, inner root sheath, and outer root sheath near the hair bulb (Fig. 5). Negative controls were presented in Supplementary Fig. 1B.
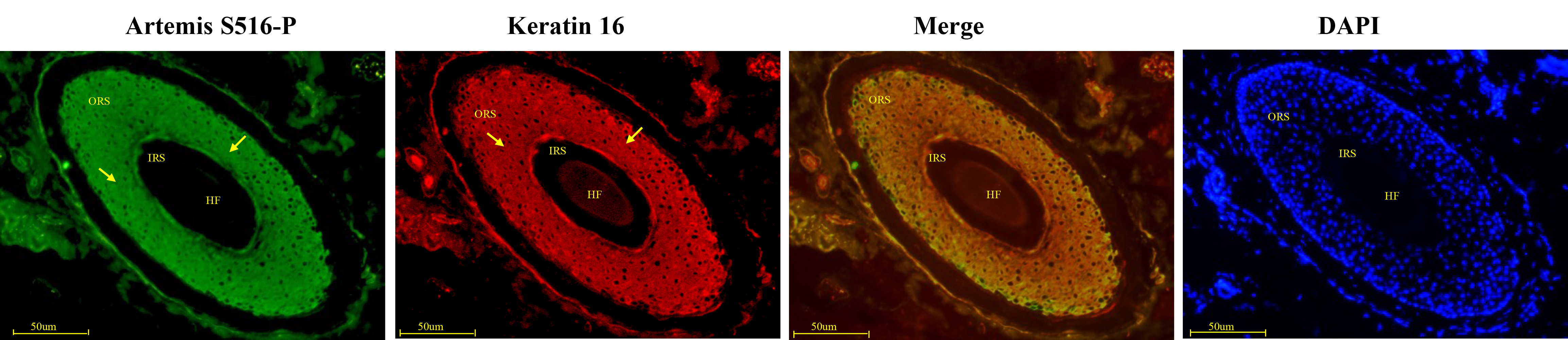 Fig. 5.
Fig. 5.Immunofluorescence double-staining for Artemis S516-P and keratin 16 in human growing scalp HF. In situ immunofluorescence double-staining of cryostat sections of scalp from healthy donors. Artemis S516-P and keratin 16 both showed strong staining patterns along the outer root sheath in the middle and upper sections of the HF, and both showed weaker staining in the hair matrix, medulla, cortex, inner root sheath, and outer hair root sheath near the hair bulb. Artemis S516-P stained green; DAPI nuclear staining was blue; keratin 16 stained red; merged staining was yellow. Positive staining was highlighted by arrows. Abbreviations: ORS, outer root sheath; IRS, inner root sheath; HF, hair follicle.
Artemis S516-P showed strong positive expression in the outer hair root sheath cells in the upper and middle sections of the HF, near the basal layer. Keratin 14 showed strong positive expression in the outer hair root sheath cells, which showed only weak expression of Artemis S516-P (Fig. 6). Negative controls were presented in Supplementary Fig. 1B.
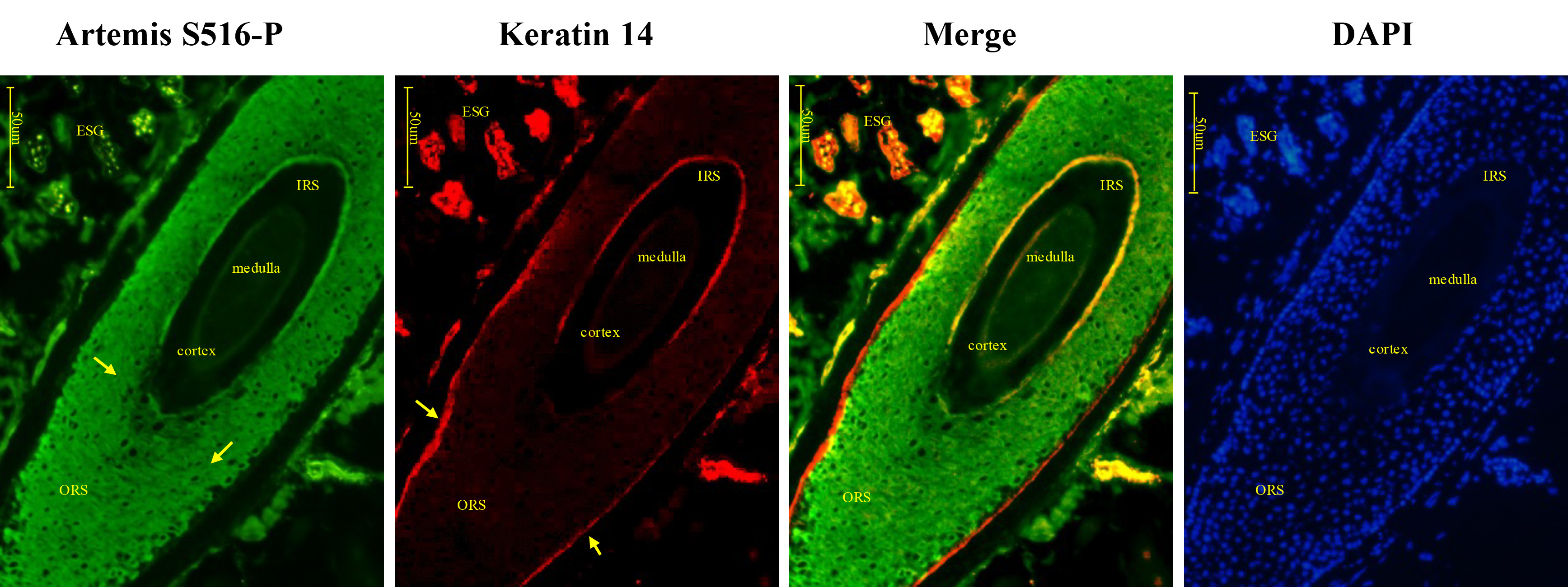 Fig. 6.
Fig. 6.Immunofluorescence double-staining for Artemis S516-P and keratin 14 in human growing scalp HF. In situ immunofluorescence double-staining of cryostat sections of scalp from healthy donors. Artemis S516-P showed strong cytoplasmic staining in the outer root sheath cells of the HF in the upper and middle sections, except in the basal layer, while keratin 14 showed strong staining in the basal layer of the outer root sheath cells, where Artemis S516-P was only weakly expressed. Artemis S516-P stained green; DAPI nuclear staining was blue; keratin 14 stained red; merged staining was yellow. Positive staining was highlighted by arrows. Abbreviations: ORS, outer root sheath; IRS, inner root sheath; ESG, eccrine sweat gland.
C-myc was expressed in the inner root sheath, close to the outer hair root sheath where Artemis S516-P was expressed. Expression of both proteins reduced near the hair bulb, indicating synchronous, rather than coincident expression (Fig. 7). Negative controls were presented in Supplementary Fig. 1C.
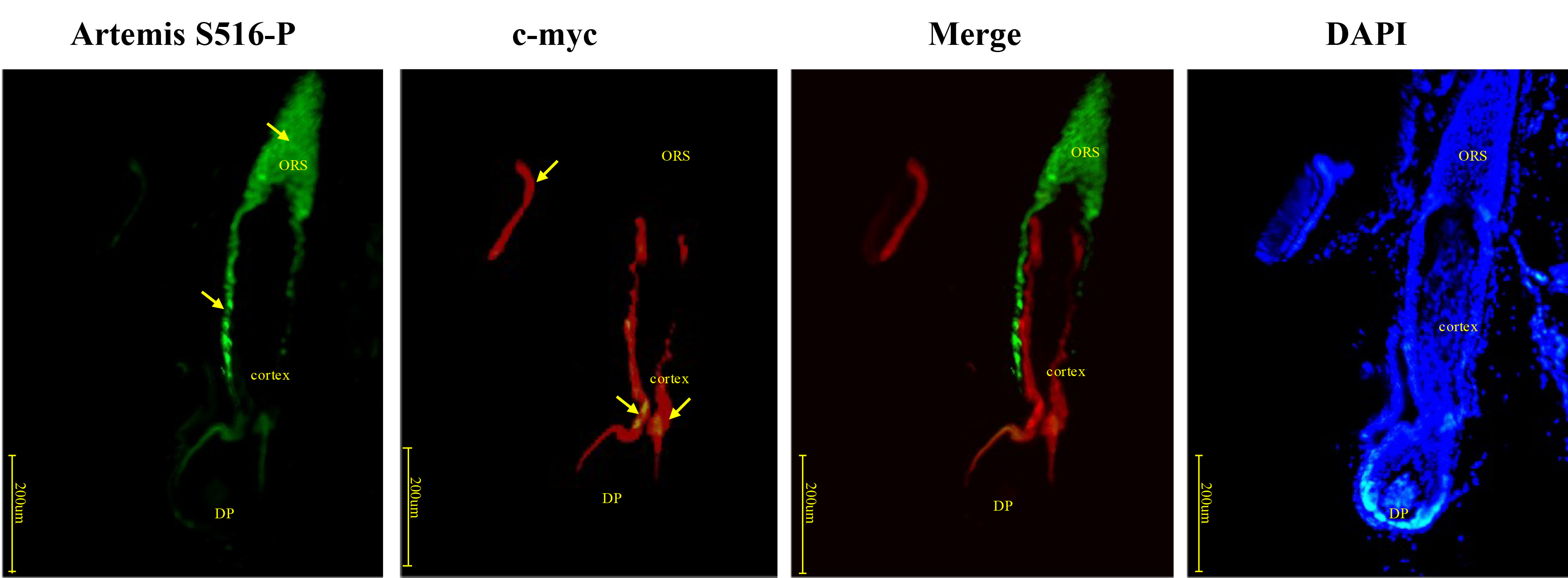 Fig. 7.
Fig. 7.Immunofluorescence double-staining for Artemis S516-P and c-myc in human growing scalp HF. In situ immunofluorescence double-staining of cryostat sections of scalp from healthy donors. C-myc was expressed in the inner root sheath, close to the outer root sheath where Artemis S516-P was expressed. Both Artemis S516-P and c-myc staining were reduced closer to the hair bulb. Artemis S516-P stained green; DAPI nuclear staining was blue; c-myc stained red; merged staining was yellow. Positive staining was highlighted by arrows. Abbreviations: ORS, outer root sheath; DP, dermal papilla.
p21 was expressed close to the cortex and medulla where Artemis S516-P was positive, while expression of both proteins became negative nearer to the hair bulb, similar to the pattern observed for c-myc (Fig. 8). Negative controls were presented in Supplementary Fig. 1C.
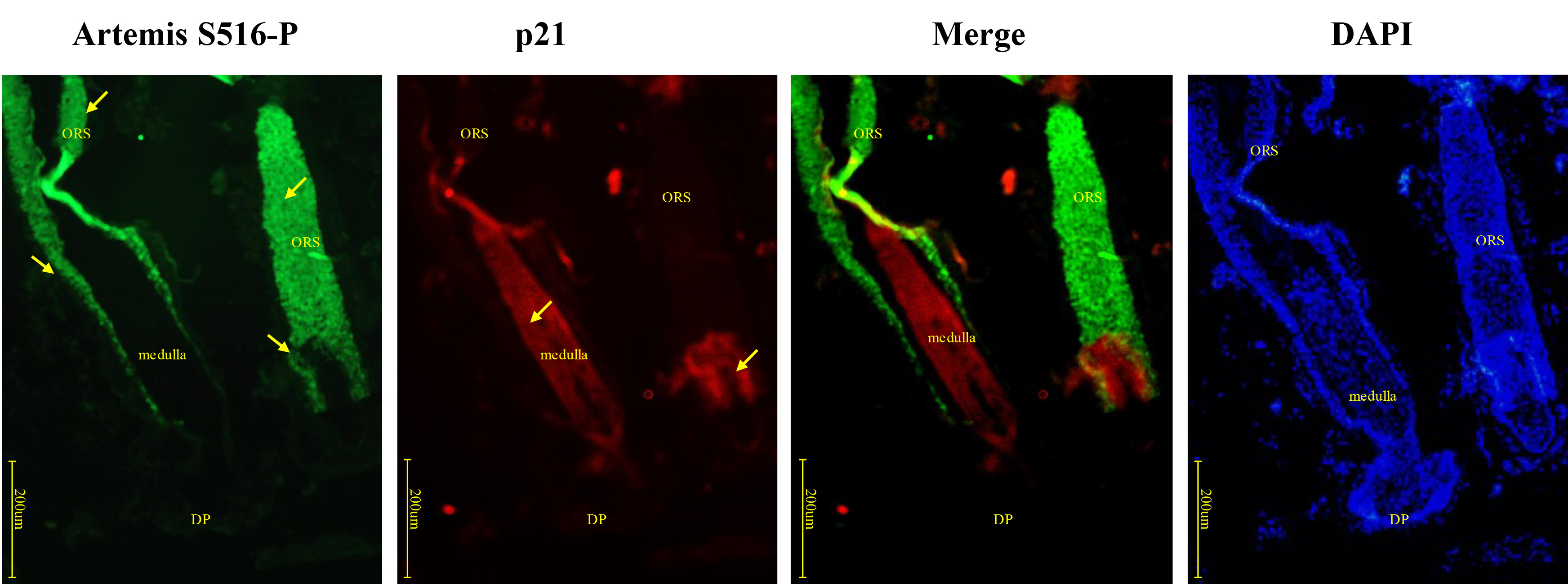 Fig. 8.
Fig. 8.Immunofluorescence double-staining for Artemis S516-P and p21 in human growing scalp HF. In situ immunofluorescence double-staining of cryostat sections of scalp from healthy donors. p21 staining was observed close to the cortex and medulla, where Artemis S516-P also showed strong staining. Both Artemis S516-P and p21 staining decreased nearer the hair bulb. Artemis S516-P stained green; DAPI nuclear staining was blue; p21 stained red; merged staining was yellow. Positive staining was highlighted by arrows. Abbreviations: ORS, outer root sheath; DP, dermal papilla.
Artemis is thought to be involved in DNA DSB repair via the NHEJ pathway [28], cell cycle checkpoint regulation [29], p53-dependent apoptosis [30], and in ubiquitin-dependent proteolysis [12]. Preliminary studies had demonstrated Artemis S516-P expression in the epidermis of the human scalp, upper inner root sheath, outer hair root sheath, hair stem progenitor cells, and hair germ cells of HF in anagen [22]. HF germ cells possess innate proliferative potential, severing as an early source for accelerating the process of HF to anagen from telogen, independent of HF bulge activity [24, 25]. In anagen HF, the cells comprising the inner root sheath as well as hair shaft proliferate and differentiate from HF germ cells, but the outer root sheath of the HF is derived from the bulge cells [24, 25]. Therefore, Artemis may regulate the process of HF progressing from telogen to early anagen. During telogen in plucking-induced hair cycling in mice, Artemis S516-P was located mainly near the club hair, germ cells and cap [23]; these regions are rich in stem cells as well as being critical in inducing new hair cycle. This suggests that Artemis S516-P may play an important role in modulating HF keratinocytes. We therefore performed this study to investigate the possible biological role of Artemis S516-P in HF, using immunofluorescence double-staining.
Here, we analyzed the co-expression patterns of Artemis S516-P with different indicators of proliferation, differentiation and apoptosis in human anagen HF. The results suggest that Artemis S516-P may have a variety of biological functions in the follicle-regulatory process. Artemis S516-P showed an opposite trend to phospho-p53 from outer hair root sheaths to inner hair root sheaths. Phospho-p53 was much intensive in hair bulb while Artemis S516-P was more intensive out of hair bulb, suggesting that Artemis S516-P may inhibit the expression of phospho-p53, thereby reducing apoptosis while promoting cell survival.
Artemis S516-P expression in human scalp anagen HF showed an opposite trend to both Bax and Bcl-2. Artemis S516-P expression decreased gradually closer to the hair bulb, while Bax and Bcl-2 expression levels increased. It is intriguing that the pro-apoptotic Bax and the anti-apoptotic Bcl-2 seem to be affected by Artemis S516-P, and this phenomenon needs further study [31, 32]. Together with its inhibition of phospho-p53, these results suggest that Artemis S516-P may be involved in regulating HF apoptosis and survival.
Notably, Artemis S516-P and keratin 10 expression levels showed opposite patterns in the inner and outer HF, and in the upper and middle sections of the HF. Artemis S516-P was expressed mainly in cells within the HF, including the upper section of the outer root sheath, while keratin 10 expression in those sections was weak; conversely, areas with strong keratin 10 expression showed weak Artemis S516-P expression. Notably, keratin 10-positive keratinocytes were located mainly outside the HF, including the epidermis, sebaceous glands and sweat glands. However, in this study, we found Artemis S516-P was expressed at a low level in those cells compared with HF. This suggests that Artemis S516-P activation may inhibit the expression of keratin 10. It was interesting that Artemis S516-P and keratin 16 were both strongly expressed in the middle and upper sections of HF with their expression patterns almost overlapped fully. Artemis S516-P may affect the expression of keratin 16. Keratin 16-positive keratinocytes were located mainly in the HF, indicating active proliferation, while keratin 10-positive keratinocytes located mainly outside the HF indicated terminal differentiation [33]. We therefore hypothesized that Artemis (serine 516) may play a decisive role in regulating the differentiation of HF pluripotent stem cells into HF, by inhibiting terminal differentiation. Artemis S516-P was expressed in the middle and upper sections of the HF, while keratin 14 was expressed strongly in the basal cells of the hair outer root sheath, where Artemis S516-P was only weakly stained. We therefore hypothesized that Artemis S516-P may play a role in regulating the differentiation of HF.
C-myc and p21 are both involved in cell cycle promotion and cell-cycle brakes impairment [34]. C-myc was expressed in the inner root sheath, while Artemis S516-P was expressed close to the outer root sheath; as Artemis S516-P expression decreased, c-myc expression also weakened in nearby tissues. Artemis S516-P may be able to affect the growth of HFs via c-myc modulation, and thus affect the growth of HF via c-myc modulation. Notably, p21 was expressed in the cortex and medulla close to Artemis S516-P -positive sites, and decreased as surrounding Artemis S516-P expression decreased. Artemis S516-P may thus regulate c-myc and p21 in similar ways.
In conclusion, our study implies the clues that Artemis S516-P may be involved in the regulation of cell proliferation, apoptosis and differentiation during HF growth and development. However, our study was a preliminary study evaluating the spatial distribution of Artemis S516-P and compared it with other molecules involved in the HF development. It is known that the expression of Artemis itself would underlie phosphorylation of S516. Therefore, the expression of Pan-Artemis should be examined and compared to Artemis S516-P. In addition, ATM and/or DNA-PKcs are thought to phosphorylate this residue in DNA damage response. In current work, we were limited in our experimental ability to address these issues on Artemis in HF. In the future, further research focusing on these issues should be carefully investigated to support this work.
Artemis, DNA cross-link repair 1C; RS-SCID, radiosensitive severe combined immunodeficiency syndrome; VDJ, variable (diversity) joining gene; IR, ionizing radiation; DSBs, DNA double-strand breaks; DNA-PKcs, DNA-dependent protein kinase catalytic subunit; ATM, ataxia telangiectasia mutated; NHEJ, non-homologous end joining; Artemis S516-P, Artemis phosphorylated at serine 516; HF, hair follicles; FITC, fluorescein isothiocyanate; DAPI, 6-diamidino-2-phenylindole segments; ORS, outer root sheath; IRS, inner root sheath; DP, dermal papilla.
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
RD and ZS designed the research study. RD performed the research. MZ and XW provided help and advice on the study design and the experiment. RD analyzed the data. RD and XW wrote the manuscript. All authors contributed to editorial changes in the manuscript. All authors read and approved the final manuscript.
This study was approved by the ethics committee of the Second Affiliated Hospital, Zhejiang University School of Medicine, China, and conformed to internationally accepted ethical standards (2012-029). All donors gave their written consent.
Not applied.
This research was funded by Natural Science Foundation of China (No. 82103754) and Zhejiang Provincial Natural Science Foundation of China (No. LQ20H110002).
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.