







 , Wenping Zhang 1,*
, Wenping Zhang 1,*1 College of Life Sciences and Medicine, Zhejiang Sci-Tech University, 310053 Hangzhou, Zhejiang, China
2 Department of Pain Medicine, Affiliated Hospital of Jiaxing University, 314000 Jiaxing, Zhejiang, China
Abstract
Lumbar sympathectomy improves blood flow to the lower limbs and is widely used in clinical practice to treat lower limb pain and cold. However, the therapeutic mechanisms underlying lumbar sympathectomy for limb coldness resulting from nerve injury remain unclear. This study aimed to investigate the effect of lumbar sympathectomy on cold allodynia in rats with spared nerve injury (SNI) and identify potential target genes associated with its analgesic effects.
A rat model of SNI was established. Mechanical and cold pain thresholds were assessed in rats with SNI to explore the analgesic effects of lumbar sympathetic neurectomy on cold allodynia. Poly(A)-seq was used to analyze the transcriptional profile of the spinal cord. Differentially expressed genes (DEGs) were screened and analyzed using bioinformatics and validated by quantitative PCR analysis.
Lumbar sympathectomy improved mechanical pain, cold allodynia, and cold sensitivity in the ipsilateral hind paw of SNI rats (all p < 0.05). Poly(A)-seq identified 278 DEGs (177 upregulated and 101 downregulated) in the spinal cords of SNI model rats compared with control rats. We identified 174 DEGs in the gene expression profile of lumbar sympathectomized SNI rats, including 69 upregulated and 105 downregulated genes, compared with SNI model rats. Functional analysis of the DEGs revealed that the most significantly enriched pathways included immune-related pathways and cellular molecular components, which mediate neuroinflammation, central sensitization, and chronic pain. To explore the correlation among the DEGs, we used the STRING database to construct protein-protein interaction networks. Finally, quantitative PCR analysis revealed six potential target genes associated with cold analgesic effects epithelial mitogen gene (EPGN), histone cluster 2 H3 family member C2 (Hist2h3c2), small integral membrane protein 6 (Smim6), family with sequence similarity 187 member a (FAM187A), LOC108349650, and LOC102550818.
Lumbar sympathectomy may alleviate cold allodynia in SNI model rats. We identified key genes associated with pain mitigation, offering potential therapeutic targets. These genes may serve as targets for treating nerve injury-induced cold allodynia. These findings provide valuable insights for the development of new treatments for nerve-related pain disorders.
Keywords
- lumbar sympathectomy
- poly(A)-seq
- cold allodynia
Neuropathic pain is triggered by damage to the nervous system and is characterized by spontaneous, abnormal pain, nociceptive hypersensitivity, and heightened sensitivity. Cold allodynia is a common symptom of neuropathic pain and manifests as nociceptive hypersensitivity to cold temperatures and pain responses to cold temperatures that would not usually cause pain [1]. Cold allodynia significantly impacts public health, as it is common in patients with neuropathic pain, including those with traumatic spinal cord injury and peripheral nerve damage. This condition severely impairs quality of life, causing physical, psychological, and emotional distress. Some patients with traumatic spinal cord injury have a low cold threshold, experiencing spontaneous and persistent cold allodynia occurring at ambient temperatures [2]. Approximately 50% of patients with spinal cord injury report that cold allodynia significantly affects their quality of life and psychological well-being [3]. Peripheral nerve injuries are a common clinical issue, with peripheral neuropathy presenting as cold allodynia experienced by approximately 2.8% of patients with polytrauma and 5% of patients with nerve root and plexus injuries [3]. The pathogenesis of neuropathic pain remains unclear, and clinical treatment for nerve injury-induced cold allodynia remains limited [4]. Current therapies are often ineffective, and patients may show resistance or insensitivity to commonly used clinical drugs. Western medicine, primarily focused on analgesia and nerve modulation methods such as transcutaneous electrical nerve stimulation and spinal cord electrical stimulation, has not significantly alleviated neuropathic pain. Given the challenges in treating cold allodynia caused by nerve damage, there is an urgent need for more effective therapeutic strategies.
The sympathetic nervous system is a component of the autonomic nervous system and plays a crucial role in maintaining numerous pain states. Local sympathetic blockade alleviates certain neuropathic pain conditions and preclinical neuropathic pain behaviors [5, 6]. Paclitaxel increases sensitivity to cold stimuli, and sympathectomy prevents the development of cold hyperalgesia in the ipsilateral hind paws of both male and female mice following paclitaxel treatment [7]. In a nerve injury model, treatment of neuropathic pain with microsympathectomy had similar effects on mechanical pain thresholds in both male and female mice [8]. In a back pain model with local inflammation, microsympathectomy had similar modulatory effects on mechanical and cold pain thresholds [9]. Clinically, lumbar sympathetic nerve block is effective for treating cold hypersensitivity in the hands and feet and improving symptoms of cold lower extremities [10]. Percutaneous radiofrequency lumbar sympathectomy can be used to treat diabetic peripheral neuropathy and type 1 complex regional pain syndrome of the lower limbs [10]. Sympathetic nerve modulation enhances vasodilation and perfusion, thereby alleviating cold sensitivity [11]. However, despite the clinical potential of sympathetic neuromodulation in the treatment of neuropathic pain, several challenges and uncertainties remain. The mechanisms underlying sympathetic neuromodulation therapy are not well understood, and the effect of local sympathectomy on the onset and progression of neuropathic cold allodynia resulting from spinal injury remains unclear.
Poly(A)-seq, a high-throughput RNA sequencing technique, has emerged as a powerful tool for profiling polyadenylated RNA transcripts. By targeting poly(A) tails, this method offers a comprehensive view of gene expression dynamics, facilitating the precise identification of key regulatory genes [12]. To investigate the mechanism through which lumbar sympathectomy alleviates cold allodynia in rats, we established a cold pain model induced by peripheral nerve injury through selective damage to sciatic nerve branches, known as the spared nerve injury (SNI) model. We performed genome-wide RNA sequencing of the spinal cord in the SNI rat model to examine alterations in the expression profiles of long non-coding RNAs and mRNAs. We investigated the key genes associated with cold sensitivity in rats following lumbar sympathectomy for reverse nerve injury. Our study provides insights into the mechanisms through which lumbar sympathectomy ameliorates cold allodynia in rats and may assist in identifying genes or signaling pathways relevant to the treatment of cold allodynia resulting from nerve injury.
In this study, we aim to explore the underlying mechanisms by identifying key genes associated with cold sensitivity following lumbar sympathectomy in nerve-injured rats using poly(A)-seq technology. By advancing the understanding of these mechanisms, our research could contribute to the development of novel therapeutic targets and improve existing treatment paradigms for cold allodynia, particularly in patients with nerve injuries.
Forty male SD (Sprague-Dawley) rats (180–200 g) were obtained from Beijing
Viton Lihua Laboratory Animal Technology Co. (Beijing, China). The rats were
housed in a strictly controlled, specific pathogen-free environment (five animals
per cage, temperature 22
SNI was used to induce cold sensitization in rats following nerve injury. After a few days of acclimatization feeding, the rats were subjected to surgical intervention, as described by Guida et al. [13], to induce cold allodynia. The SNI model was established by deeply anesthetizing the animal through an intraperitoneal injection of sodium pentobarbital (P3761, Sigma-Aldrich, Darmstadt, Germany) at a concentration of 35 mg/kg and shaving the right side below the pelvis, followed by the removal of skin and muscle of the hind limbs to expose the sciatic nerve and its terminal branches. The peroneal and tibial nerves were isolated from the fascia and transected distally after being ligated with a No. 5.0 silk thread. Incisions in the muscle and skin layers were closed using sutures. All surgeries were conducted under aseptic conditions. Following surgery, the rats were placed on a warm blanket and returned to their cages after recovering from anesthesia [14]. The success rate of the SNI model typically ranges from 70% to 90%. In this experiment, the successfully established SNI model rats were expected to exhibit pain-related behavioral responses, along with changes in body weight and reduced activity.
After the rats were deeply anesthetized with an intraperitoneal injection of sodium pentobarbital at a concentration of 35 mg/kg, they were positioned in the supine position. The rat’s abdominal area was shaved and disinfected with povidone-iodine, ensuring a sterile operating environment. A longitudinal incision of approximately 3–4 cm was made along the lower segment of the rat’s abdominal midline. The skin and subcutaneous tissues were carefully separated, avoiding damage to deeper organs. A longitudinal incision of approximately 3–4 cm was made in the lower midline of the rat’s abdomen. An abdominal retractor was used to expose the abdominal cavity, carefully separating the skin and subcutaneous tissue to avoid damage to deep organs. Landmarks such as the right kidney, inferior vena cava, and iliolumbar veins were identified. Dissect the fascia on the right side of the inferior vena cava to expose the lumbar sympathetic nerve adjacent to the abdominal aorta. Using fine forceps and microsurgical scissors, carefully excise the unilateral lumbar sympathetic chain, including the sympathetic ganglia from L2 to L4, while minimizing damage to nearby blood vessels and tissues. Postoperatively, absorbable sutures were used to close the abdominal muscles and skin layer by layer, ensuring the incision remained tension-free and minimizing the risk of infection.
The normal group (N) rats serve as the control group and undergo no nerve injury or surgical interventions. They are subjected only to standard acclimatization feeding and regular management. The SNI model group (S) rats undergo only the SNI surgical procedure. The lumbar sympathectomy normal group (NL) rats undergo only lumbar sympathectomy without the SNI procedure. The SNI-followed lumbar sympathectomy group (SL) rats first undergo the SNI procedure, followed by lumbar sympathectomy.
Before performing the behavioral study, the rats were individually placed in a transparent Plexiglas chamber on a raised mesh floor for 30 minutes to acclimate them to the environment. An electronic pain meter (Almemo 2450, AHLBORN, Holzkirchen, The Netherlands) was used to measure changes in the mechanical pain threshold (paw withdrawal threshold, in g) in different groups of rats. During the test, a wire probe was used to apply force vertically to the mid-plantar surface of the right hind paw of the rats; sudden retraction, licking, or violent shaking of the hind paw in response to the stimulus was considered a pain-like reaction and the pain threshold was recorded. The average paw withdrawal threshold was determined from every five measurements taken at 30-second intervals.
Cold pain and cold sensitivity in the rats were measured using a hot and cold plate analyzer (Beijing Zongshi Dichuang Science and Technology Development Co., Ltd., Beijing, China). The rats were placed in the experimental environment for 15–30 minutes to minimize interference from environmental factors. Changes in cold and hot pain over time in each group were assessed by recording the number of times the rats lifted their feet at 4 °C, with measurements repeated three times for each rat. The minimum tolerated temperatures of rats in each group were determined by downward gradient cooling, with 37 °C used as a benchmark, and the measurements were repeated three times for each rat.
Observe the rat’s response to acetone (Sigma-Aldrich, Catalog No. 650501) stimulation, which typically manifests as frequent lifting of the foot, licking, or scratching. These behaviors are associated with the rat’s sensitivity to cold stimuli, with stronger reactions indicating higher cold pain sensitivity. The number of lifts within 1 minute was recorded to assess the rat’s cold sensitivity, with measurements repeated three times for each rat.
Rats in the N, S, and SL groups were deeply anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneally) on day 21, followed by myocardial perfusion with 0.9% saline (4 °C). After perfusion, segments L4–L6 of the dorsal horn of the spinal cord were harvested. Total RNA was extracted from rat spinal cord tissues using a Magzol kit (Magen Biotech, Catalog No. R480101, Guangzhou, China), and RNA yield and integrity were assessed using a K5500 microspectrophotometer (Beijing Kaiao, Beijing, China) and an Agilent 2200 TapeStation (Agilent Technologies, Santa Clara, CA, USA), respectively. The mRNA was enriched using oligo (dT) according to the instructions of NEBNext® Poly(A) mRNA Magnetic Isolation Module (New England Biolabs, Ipswich, MA, USA). Subsequently, the mRNA was fragmented to approximately 200 bp.
The RNA fragments underwent first- and second-strand cDNA synthesis, followed by junction ligation and low-cycle enrichment, according to the instructions of the NEBNext Ultra RNA Library Preparation Kit (New England Biolabs, Catalog No. E7530S, Ipswich, MA, USA). The purified library products were evaluated using an Agilent 2200 TapeStation and Qubit (Thermo Fisher Scientific, Catalog No. Q32852, Waltham, MA, USA), and the libraries were prepared using Illumina (San Diego, CA, USA) at RiboBio Co., Ltd. (Guangzhou, China) with a 150 bp paired-end approach for sequencing.
Raw FASTQ sequence data were processed with Trimmomatic software (version 0.36,
https://github.com/usadellab/Trimmomatic). Specifically, end sequences with Phred
quality scores below 20 were removed (software options: TRAILING, 20; SLIDING
WINDOW, 4:15; MINLEN, 52) to standardize sequence lengths for the downstream
clustering process. The paired-end reads were aligned to the rat reference genome
(rn6) using HISAT2 (The University of Maryland, College Park, MD, USA). HTSeq
(version 0.12.4, https://github.com/simon-anders/HTSeq) was used to count the
reads mapped to each gene. Statistically significant DEGs were obtained using
DESeq2 software (version 1.40.2)
(https://bioconductor.org/packages/release/bioc/html/DESeq2.html) (by adjusting
for p-values
DEGs were analyzed for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment using the “clusterProfiler” package (https://bioconductor.org/packages/release/bioc/html/clusterProfiler.html) in R Bioconductor software (version 3.19) (https://bioconductor.org/). To explore the correlation between these DEGs, protein-protein interaction networks were constructed using the STRING database (https://cn.string-db.org/) and visualized using Cytoscape software (version 3.10.1) (https://cytoscape.org/).
Six DEGs (four genes with upregulated expression according to transcriptomic analysis: small integral membrane protein 6 (Smim6), LOC102550818, LOC108349650, and family with sequence similarity 187 member a (FAM187A); and two genes with downregulated expression, epithelial mitogen gene (EPGN), and histone cluster 2 H3 family member C2 (Hist2h3c2)) were selected for quantitative reverse transcription polymerase chain reaction (qRT-PCR) validation. Total RNA was extracted from rapidly frozen spinal cord tissue using TRIzol reagent (Thermo Fisher Scientific, Catalog No. 15596026). RNA quality and concentration were determined using a NanoDrop-2000 spectrophotometer (Thermo Fisher Scientific). Reverse transcription was performed using SweScript All-in-One RT SuperMix for qPCR (One-Step cDNA Remover) (ServiceBio, Catalog No. G3330, Wuhan, China). The target gene mRNA was quantified using qRT-PCR with 2X SG Fast qPCR Master Mix (B639271, BBI, Roche, Basel, Switzerland) and 2.5 µM gene primers. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used for normalization. The Transcript Data®-Clark method was calculated using the 2-ΔΔCt method to determine the expression levels of target genes relative to that of GAPDH. The primer details used for qRT-PCR validation are presented in Table 1.
| Gene name | Primer sequence (5′-3′) | Product size |
| GAPDH-F | CAAGTTCAACGGCACAGTCAA | 140 bp |
| GAPDH-R | CGCCAGTAGACTCCACGACA | |
| Hist2h3c2-F | GATCCGGCGCTACCAGAAG | 170 bp |
| Hist2h3c2-R | TGTCCTCGAACAGACCCACC | |
| Smim6-F | AAGACCATCCAGAATGAAGCC | 156 bp |
| Smim6-R | TTCACGCACATCCTTTCTCC | |
| LOC102550818-F | TTTTCCCTCCTGGATTCTTCA | 104 bp |
| LOC102550818-R | ACGGCTTGATATGCCCTTAAC | |
| LOC108349650-F | CTTTTGAGTCGGGGAGAACTATGTG | 170 bp |
| LOC108349650-R | CCGCATATTGTTCTAATCAGTCC | |
| FAM187A-F | CGAATGTTCAGCCTGTTGGT | 214 bp |
| FAM187A-R | GGGTCCACTCCCAGAAGGTA | |
| EPGN-F | TTTGGGAGTTCCAATATCAGC | 112 bp |
| EPGN-R | TGTGATTGGAGGTGTTACAGTCA |
FAM187A, family with sequence similarity 187 member a; Smim6, small integral membrane protein 6; EPGN, epithelial mitogen gene; Hist2h3c2, histone cluster 2 H3 family member C2.
Data are presented as means
We established a rat model of SNI by surgically inducing selective injury of the
sciatic nerve branches. Lumbar sympathetic nerve resection was performed on day 5
post-SNI surgery for both the SL and NL groups. Mechanical pain thresholds in the
hind paw and hot and cold pain sensitivity tests were conducted on the day before
the SNI surgery (pre) and post-surgery days 2, 5, 8, 11, 13, 17, and 20. Notably,
a lumbar sympathectomy was performed on SNI rats on day 6. We illustrated the
timeline of the experimental procedure, showing the time points for the different
behavioral tests and the surgical interventions (SNI surgery and lumbar
sympathectomy) (Fig. 1A). We observed that, compared to the N group, the S group
exhibited significantly lower mechanical pain thresholds (p
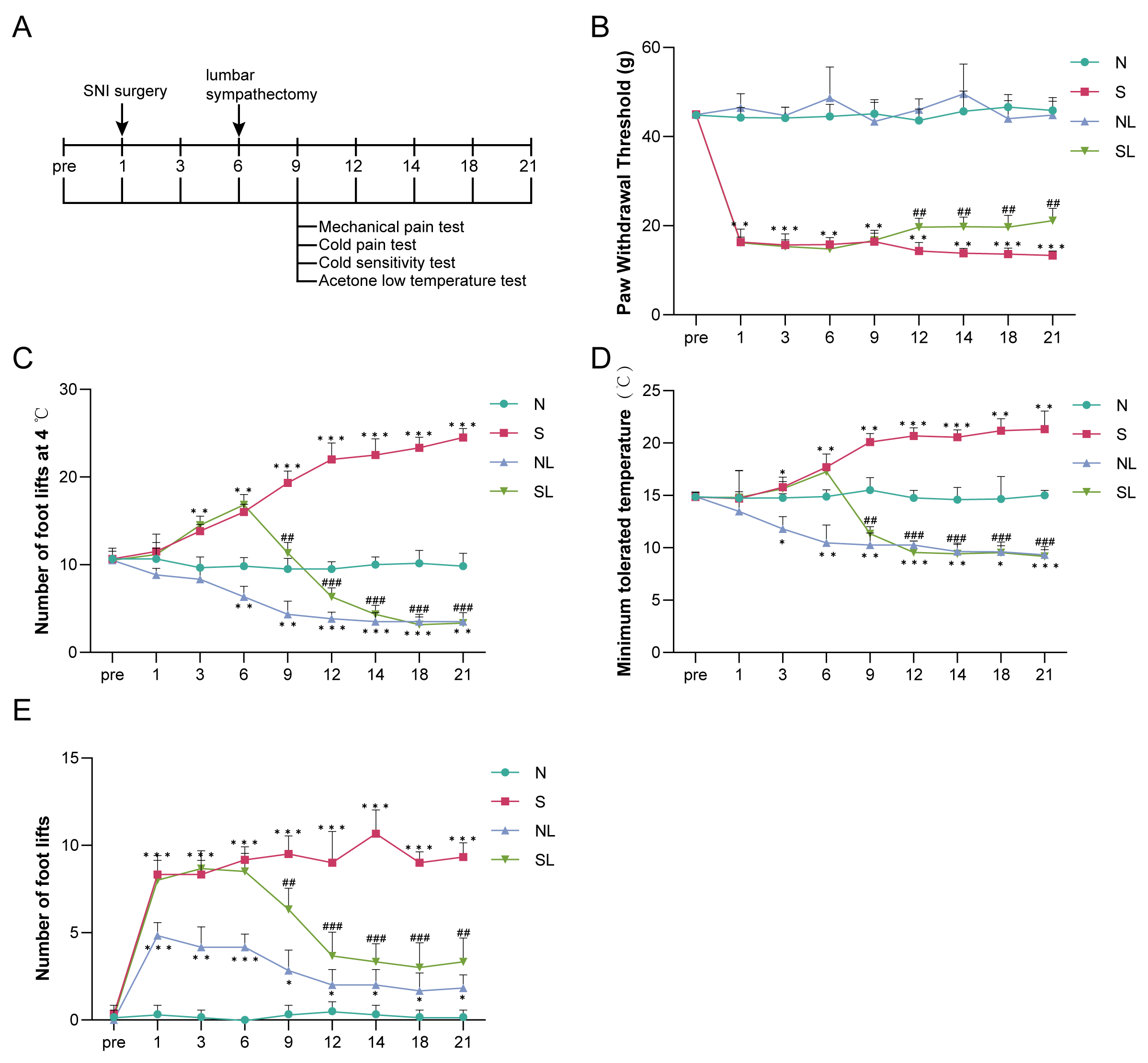 Fig. 1.
Fig. 1.
Lumbar sympathectomy attenuates abnormal pain induced by nerve
injury in spared nerve injury (SNI) model rats. (A) Establishment of a rat model
of SNI and experimental protocol for performing lumbar sympathectomy. (B)
Mechanical withdrawal threshold (MWT). (C) Cold pain sensitivity. (D) Cold
allodynia. (E) Acetone. Compared with the normal group (N), *p
In addition, compared with the normal group, the lumbar sympathetic nerve of
normal rats was removed without affecting their perception of mechanical
stimulation (p
To analyze the neurophysiological mechanisms involved in lumbar sympathectomy modulating cold sensitivity in the SNI rat model, we collected spinal cord segments L4–L6 from normal rats (N), SNI model rats (S), and lumbar sympathectomized SNI model rats (SL), and analyzed the transcriptional profiles of their spinal cords using poly(A)-seq. Each sequencing sample yielded approximately 50.3 million, 69.1 million, and 64.4 million raw reads in the N, S, and SL groups, respectively, with the average valid reads accounting for approximately 96.86%, 96.30%, and 96.78%, respectively. More than 94.90%, 94.99%, and 94.81% of the valid reads mapped to the rat genome, respectively (Table 2). The saturation curve increased with the number of reads and then flattened (Fig. 2A). The coverage homogeneity curve indicated that the transcripts covered the rat genome homogenously (Fig. 2C), confirming that a sufficient amount of data was detected. We performed hierarchical clustering and principal component analysis to evaluate changes in the spinal cord expression profiles among sample groups based on the transcripts per million data of all expressed genes obtained through sequencing. First, hierarchical clustering showed that the samples of rats in group S had high similarity and differed from those of rats in groups N and SL (Fig. 2B). In addition, principal component analysis showed that the expression profiles of rats in group S differed from those of rats in groups N and SL. In contrast, the spinal cord expression profiles of rats in groups N and SL partially overlapped (Fig. 3A). These results suggest that spinal cord gene expression was altered in SNI model rats compared with the normal group and that lumbar sympathectomy shifts the expression profile of model rats closer to that of normal rats.
| Sample | Total raw reads | Total clean reads | Total mapped | Clean rate (%) | Clean reads Q30 (%) | Total mapping ratio (%) |
|---|---|---|---|---|---|---|
| N1 | 53,553,874 | 51,840,096 | 49,143,719 | 96.80 | 94.15 | 94.80% |
| N2 | 51,193,400 | 49,522,964 | 46,942,822 | 96.74 | 94.27 | 94.79% |
| N3 | 45,982,496 | 44,618,464 | 42,440,762 | 97.03 | 94.51 | 95.12% |
| S1 | 77,817,878 | 74,974,732 | 71,308,382 | 96.35 | 93.98 | 95.11% |
| S2 | 57,842,858 | 55,675,272 | 52,909,084 | 96.25 | 93.78 | 95.03% |
| S3 | 71,621,474 | 68,971,762 | 65,418,894 | 96.30 | 93.63 | 94.85% |
| SL1 | 68,767,702 | 66,441,212 | 62,953,703 | 96.62 | 94.14 | 94.75% |
| SL2 | 68,153,930 | 66,078,474 | 62,762,749 | 96.95 | 94.33 | 94.98% |
| SL3 | 56,126,520 | 54,313,354 | 51,440,859 | 96.77 | 94.19 | 94.71% |
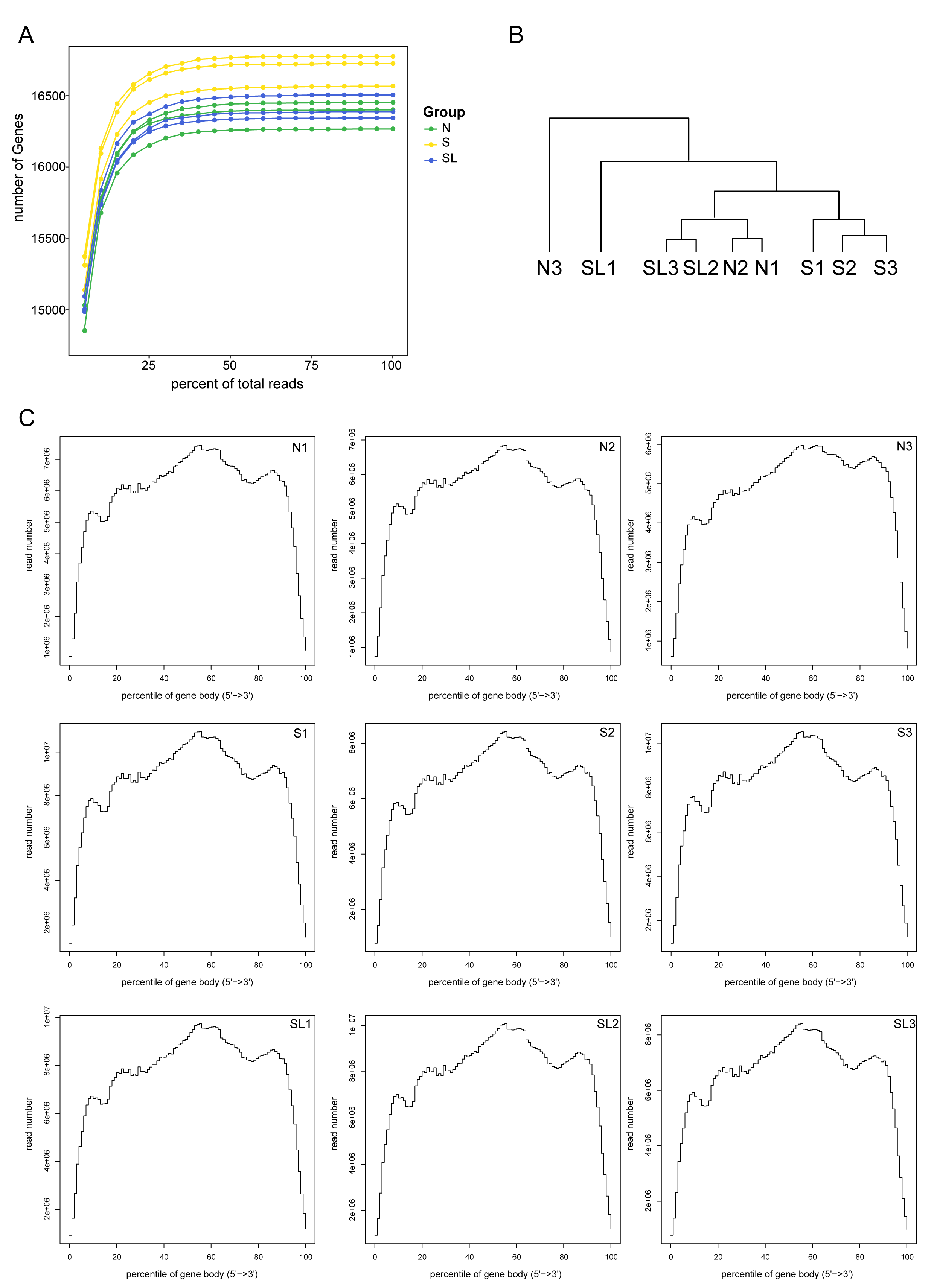 Fig. 2.
Fig. 2.
Quality assessment of rat spinal cord tissue using poly(A)-seq. (A) Sequencing saturation results of each sample. (B) Hierarchical cluster analysis dendrogram for each sample. (C) Transcriptome gene sequencing frequency and gene location distribution of each sample.
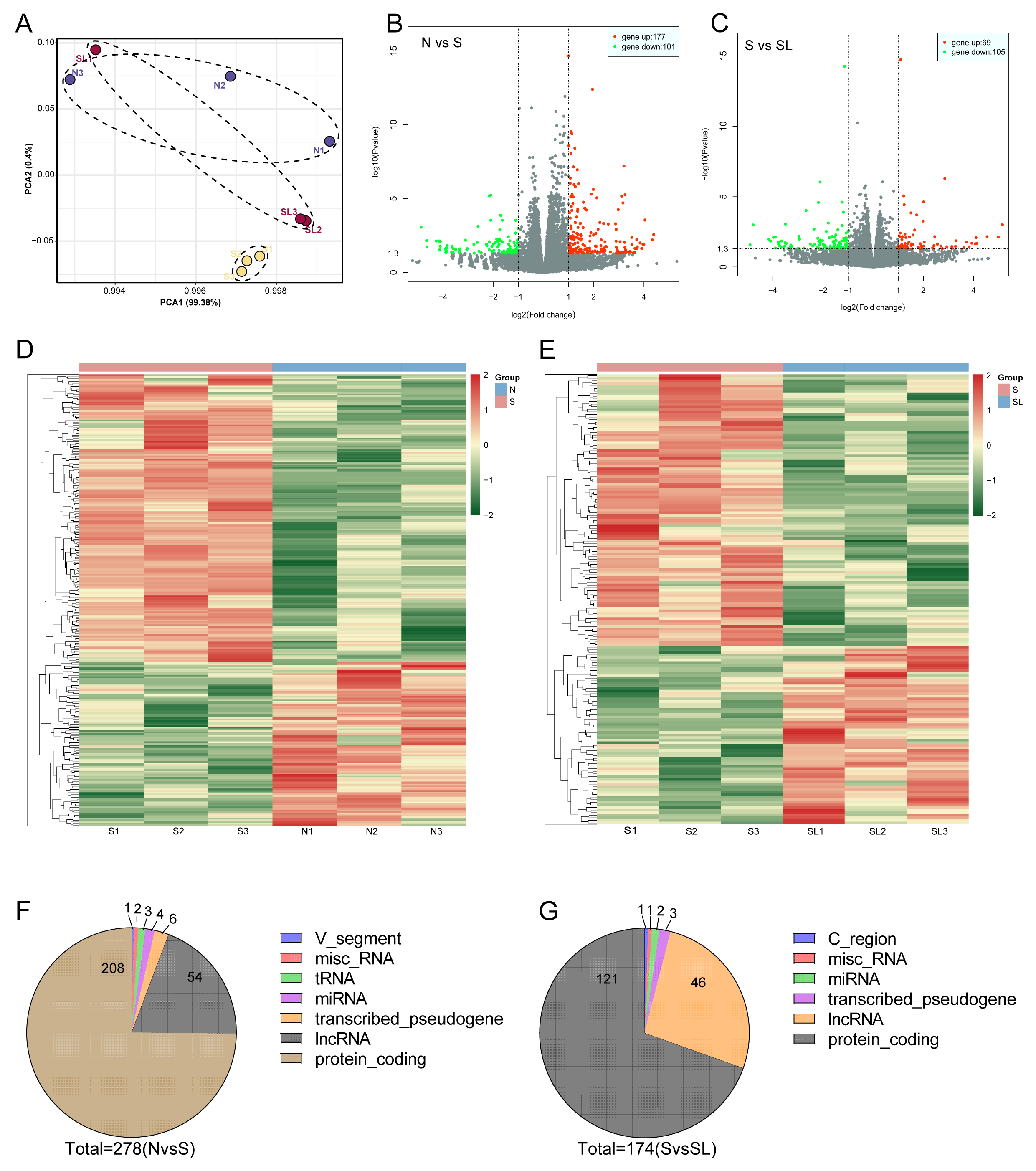 Fig. 3.
Fig. 3.
Lumbar sympathectomy modulates differentially expressed genes in the spinal cord of SNI rat. (A) Principal component analysis of spinal cord expression profiles of normal rats (N), SNI model rats (S), and lumbar sympathectomized SNI model rats (SL). (B) Volcano plot of differentially expressed genes (DEGs) in the spinal cord tissues of rats in groups S and N. Red dots indicate upregulated DEGs, green dots indicate downregulated DEGs and grey dots indicate non-significant differences in DEGs. (C) Volcano plot of DEGs in spinal cord tissues of rats in groups S and SL. Red, green, and grey dots have the same meanings as those in (C). (D) Heatmap inset of hierarchical clustering analysis of DEGs in the spinal cord tissues of rats in groups S and N. (E) Heatmap inset of hierarchical clustering analysis of DEGs in the spinal cord tissues of rats in groups S and SL. (F) Genotype classification statistics of DEGs in the spinal cord tissues of rats in groups S and N. (G) Genotype classification statistics of DEGs in the spinal cord tissues of rats in groups S and SL. PCA, principal component analysis; miscRNA, miscellaneous RNA; tRNA, transfer RNA; miRNA, micro RNA; lncRNA,long non-coding RNA.
We screened for DEGs with log2 (fold-change)
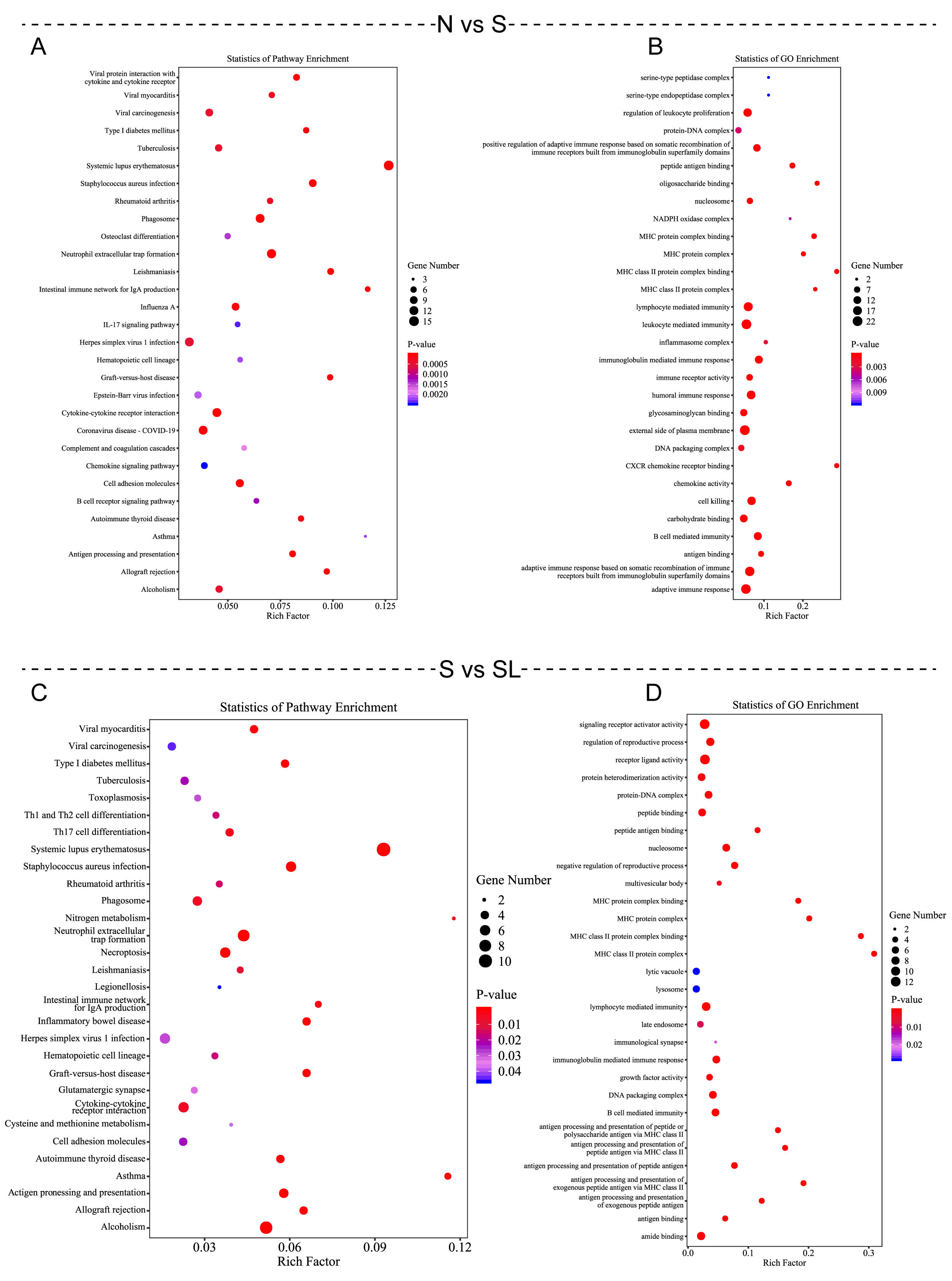 Fig. 4.
Fig. 4.
Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) enrichment were performed to analyze differentially expressed mRNAs. (A,C) KEGG enrichment bubble chart showing common DEGs. (B,D) GO enrichment bubble chart showing common DEGs.
Using Venn plot analysis, we identified 46 intersecting DEGs between SNI model
rats and SNI rats after lumbar sympathectomy (Fig. 5A). We
constructed a protein-protein interaction network of the intersecting genes using
STRING software (https://cn.string-db.org/) and found that 26 genes were involved
in protein interactions, including guanine nucleotide-binding protein
g(4) subunit (Gng4), C-X-C motif chemokine ligand 10 (also known as IP-10)
(Cxcl10), Cluster of differentiation 74 (also known as
invariant chain) (Cd74), and C-X-C chemokine receptor type 3(Cxcr3) (Fig. 5B). We used a heatmap to display the expression patterns
of selected lncRNAs and mRNAs in three experimental groups (N, S, and SL), with
hierarchical clustering distinguishing lncRNA (upper section) and mRNA (lower
section). Some lncRNAs (LOC1025508018, LOC108349650) and mRNAs
(FAM187A, Smim6, etc.) were upregulated in the S group, represented by
red or yellow in the color bar, while some genes (EPGN,
Hist2h3c2, etc.) were downregulated in the S group, shown in blue in the
color bar, and returned to levels close to those in the N group in the SL group.
This result suggests that these genes may play an essential role in the process
of cold allodynia induced by nerve injury. The bar plot on the right shows the
statistical significance of the differential expression of various genes, with
the red dashed line indicating the p-value threshold (0.05). A total of
19 genes were statistically significant (p
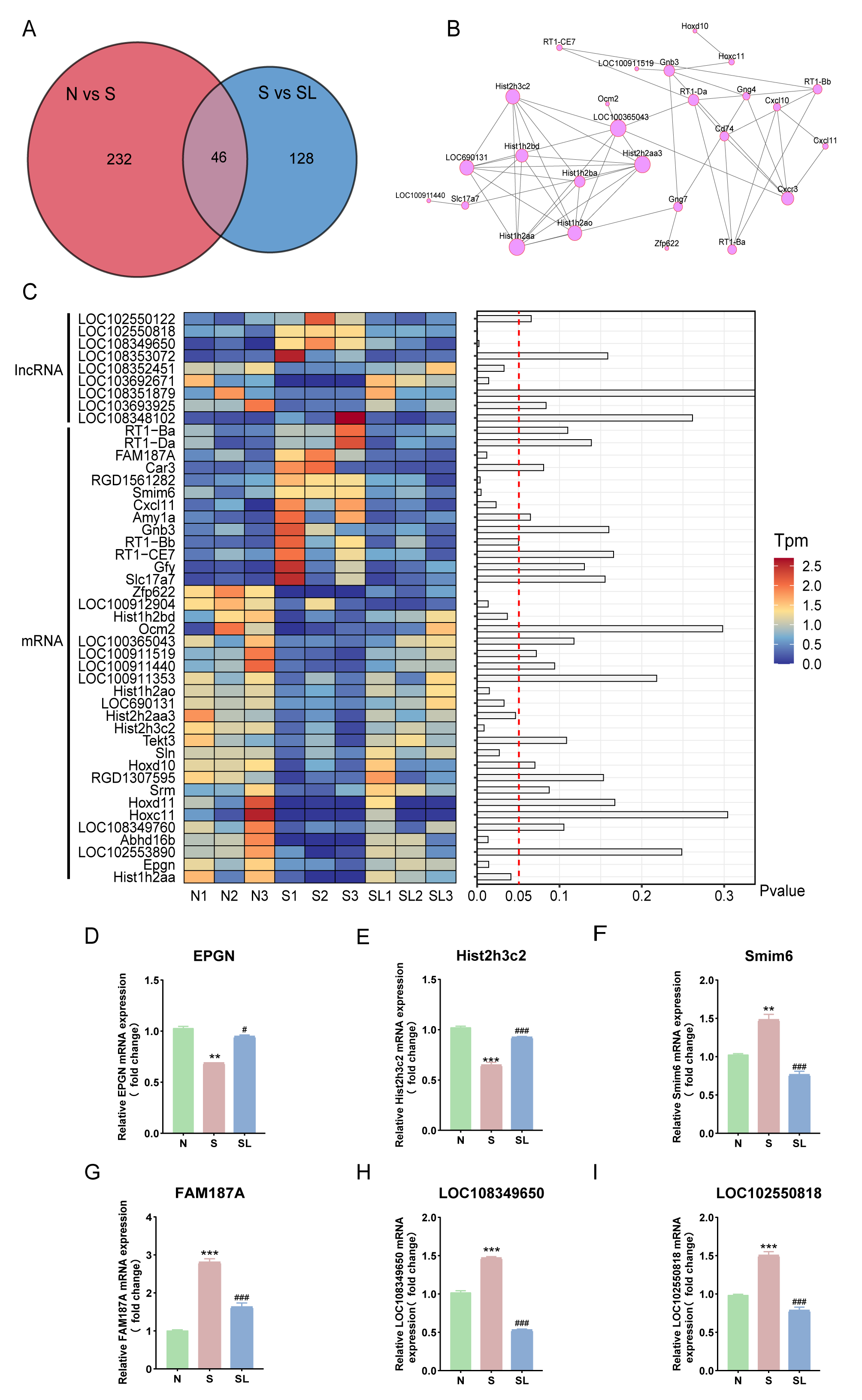 Fig. 5.
Fig. 5.
Screening and identification of core genes responsible for the
effect of lumbar sympathectomy in improving limb cold allodynia caused by nerve
injury. (A) Venn diagram of intersecting genes of two sets of DEGs (N vs. S, S
vs. SL). (B) Protein-protein interaction network constructed from common DEGs.
Grey lines indicate interactions between the corresponding proteins of a gene; a
larger circle indicates that the gene is important. (C) A
heatmap and bar plot analysis of differentially expressed lncRNAs and mRNAs
across three experimental groups (N, S, and SL) with three biological replicates
each. Relative mRNA expression of (D) EPGN, (E) Hist2h3C2, (F)
Smim6, (G) FAM187A, (H) LOC108349650, and (I)
LOC102550818. Compared with group N, **p
Nociceptive hypersensitivity to innocuous cold (or cold allodynia) is a common symptom of neuropathic and inflammatory pain caused by peripheral nerve injury [15]. Following nerve injury, neuropathic pain can arise due to atypical connections or misinterpretation of peripheral axons by the spinal cord, enhancing the integration of sensory afferent nerves into spinal cord circuitry [14, 16]. Several central mechanisms may lead to hypersensitivity to mechanical/thermal stimuli, abnormal cold nociception, and nociceptive hypersensitivity after traumatic nerve injury [17, 18], which can significantly impact the patient’s quality of life. Currently, no effective treatment exists for this hypersensitivity. In this study, we investigated cold hyperalgesia, cold intolerance, and cold sensitivity resulting from nerve injuries. Cold intolerance is believed to result from a disorder of the vascular system or neuromodulation, leading to impaired thermoregulation of the limbs and skin or altered neural responses to cold stimuli. Patients with post-traumatic cold intolerance have dysfunctional tiny nerve fiber endings, which influence chronic neuropathic pain syndrome [19]. Unfortunately, no standard treatment exists for this condition. Apart from avoiding exposure of the hands to cold stimuli and wearing protective clothing, including heated gloves, cold intolerance is closely related to the extent of sensory recovery [20]. Therefore, various rehabilitation techniques should be considered to improve sensory recovery. Injection of botulinum toxin type A around the vascular bundles of the metacarpophalangeal nerves on both sides of the affected finger, producing a temporary sympathetic blockade, can improve the symptoms of cold intolerance in post-traumatic limb patients [21]. However, this method is still underdeveloped, and further research, along with real-world analysis, is required. Nerve grafting has been a routine practice for treating traumatic peripheral nerve injury since the 1960s [22, 23, 24]. This method not only alters peripheral signaling pathways but also stimulates plastic changes in the central nervous system, which are essential for the recovery of sensory function [25, 26]. Spinal cord injuries typically result in irreversible loss of sensory and motor functions below the level of injury. Nerve grafting has emerged as a key technique for restoring sensory temperature normalization in patients with spinal cord injury. However, nerve grafting to reestablish lower limb function in patients with spinal cord injury remains a significant challenge. In addition, previous clinical study indicates that successful blockade of the sympathetic nerves leads to vasodilation, increased blood flow to the corresponding areas, and, consequently, an increase in body temperature [27]. A lumbar sympathectomy plays a crucial role in treating neuropathic pain [28]. A lumbar sympathetic nerve block involves sympathetic afferents and provides pain relief while improving blood flow to the lower extremities and increasing skin and limb temperature [29].
However, the therapeutic mechanism of lumbar sympathetic neurectomy for cold sensitivity induced by peripheral nerve injury remains unclear despite its widespread use in studying neuropathic pain [30, 31]. Therefore, in this study, an SNI model was used to induce cold allodynia in rats, which involves less tissue damage, more intuitive and more straightforward experimental steps, better behavioral reproducibility, and shows clinical manifestations similar to those of neuropathic pain [32, 33]. We confirmed that the mechanical pain threshold and cold allodynia thresholds in the SNI group were significantly lower than those in the normal group, confirming that we successfully established an SNI model for inducing cold allodynia. We tested the effects of lumbar sympathectomy on SNI-induced cold allodynia. The success of the sympathectomy technique is typically assessed by an increase in the temperature of the ipsilateral limb relative to that of the contralateral limb and by a reduction in pain duration [34]. In addition, increased tolerance to cold sensations can serve as an assessment parameter [35].
In this study, we explored the therapeutic potential of lumbar sympathectomy in alleviating cold allodynia in a rat model of SNI. The SNI model is widely recognized for its ability to mimic the clinical manifestations of neuropathic pain, making it an ideal model for studying the mechanisms underlying cold sensitivity. Our results demonstrated that lumbar sympathectomy significantly improved mechanical pain thresholds and reduced cold sensitivity in SNI rats, suggesting its potential role in modulating pain-related behaviors. This finding suggests the potential efficacy of sympathetic nerve blocks in managing neuropathic pain conditions.
The molecular mechanisms underlying the beneficial effects of lumbar sympathectomy were further investigated using poly(A)-seq, a high-throughput RNA sequencing method. This approach allowed us to profile the transcriptome of the spinal cord in SNI rats and identify DEGs related to cold sensitivity. Our analysis revealed significant changes in gene expression patterns, with notable enrichment in immune-related pathways and cellular components. This finding suggests that neuroinflammation and immune responses play a crucial role in the development and maintenance of cold allodynia. By modulating these pathways, lumbar sympathectomy may effectively reduce pain hypersensitivity and enhance overall pain management.
One of the key findings of this study was the identification of specific genes, such as Smim6 and EPGN, that showed significant differential expression in response to lumbar sympathectomy. Smim6, an endoregulatory protein, interacts with the sarcoplasmic/endoplasmic reticulum calcium pump (SERCA), a critical regulator of calcium homeostasis in neurons [36]. Dysregulation of Smim6 and SERCA has been implicated in neuropathic pain conditions, making them potential therapeutic targets [37, 38]. EPGN, a ligand for the epidermal growth factor receptor (EGFR), is involved in neurotrophic pain and has been shown to mediate hypersensitivity in various animal pain models [39, 40, 41]. Targeting the EPGN-EGFR pathway may provide new strategies for managing neuropathic cold pain.
While our study provides valuable insights into the mechanisms of lumbar sympathectomy in alleviating cold allodynia, several limitations must be acknowledged. The SNI model, although effective for studying neuropathic pain in rats, may not fully replicate the complexity of human cold allodynia. Future research should focus on translating these findings to clinical settings, with particular emphasis on understanding how these molecular pathways operate in human patients. Additionally, the relatively small sample size in this study limits the generalizability of the results, and larger cohorts are needed to confirm the reproducibility of our findings.
In conclusion, this study highlights the therapeutic potential of lumbar sympathectomy in managing cold allodynia and identifies key molecular targets involved in its mechanism. Future research should aim to validate these findings in clinical settings and explore the development of targeted therapies based on the identified pathways, with the ultimate goal of improving the quality of life for patients suffering from neuropathic cold pain.
Our findings suggest that lumbar sympathectomy attenuates cold allodynia and increases mechanical pain thresholds in rats with SNI. We also identified DEGs in the spinal cord of SNI-conditioned rats, which may serve as potential targets for treating cold allodynia.
The datasets analyzed during the current study are not publicly available due to privacy concerns. However, they are available from the corresponding author upon reasonable request.
XM designed the research methods and experimental design, conducted data collection, processing, and analysis, wrote the first draft of the manuscript and participated in discussions and revisions. HRF, PX, and LY participated in the research design and method selection, assisted in data collection, processing, and analysis, contributed to writing and revising the manuscript. YF and WPZ were responsible for the overall planning, design, and implementation of the research. They had oversight and coordination responsibility for the entire research process. Ensured the integrity and accuracy of the research. Wrote and revised the manuscript. Provided financial and technical support. All authors read and approved the final manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work.
This study and included experimental procedures were approved by the institutional animal care and use committee of Jiaxing University Medical College (JUMC2023-082). All animal housing and experiments were conducted in strict accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Not applicable.
This research received no external funding.
The authors declare no conflict of interest.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.