1. Introduction
Alzheimer’s disease (AD) is a chronic and progressive neurodegenerative disease.
Extensive distribution of -amyloid (A) plaques and
neurofibrillary tangles (NFTs) in the brain are the major histopathological
hallmarks of AD [1, 2]. One of the hypotheses of AD pathogenesis suggests that
amyloid plaques or A-peptides act as the direct cause and initiate
cascade effects, leading to a series of reactions such as tau pathology,
inflammation, synaptic dysfunction, neuronal loss, and ultimately dementia [2].
AD neuropathology initiated by A deposition is believed to begin
decades prior to the onset of clinic symptoms, and A deposition has been
observed in the cortex at 6 weeks of age and in the hippocampus at 3–4 months of
age in Amyloid precursor protein/Presenilin 1 (APP/PS1) AD mouse models [2, 3]. Hyperactivity of glycogen
synthase kinase-3 (GSK3) is believed to be critically involved in the enhancement
of A from -amyloid precursor protein (APP) and in tau
hyperphosphorylation [4, 5]. GSK3 is a highly conserved protein kinase for
glycogen synthase and is encoded by two highly related genes: GSK3
(51 kDa) and GSK3 (47 kDa). Under
physiological conditions, GSK3 is activated via phosphorylation at tyrosine
residue 279 of GSK3 (p-GSK3 tyr279) and tyrosine residue 216
of GSK3 (p-GSK3 tyr216) and inhibited via phosphorylation at
serine residue 21 of GSK3 (p-GSK3 ser21) and serine residue 9
of GSK3 (p-GSK3 ser9) [6]. The neuronal reaction to A
and tau proteopathic stress is complex, and whether the GSK3 biochemical response
is similar in the cortex and hippocampus is largely unknown.
Given that GSK3 is relevant to the pathophysiological processes of AD,
inhibition of GSK3 activity has been deemed to be an optional strategy in AD
prevention and treatment. Physical inactivity is generally believed to be one of
the risk factors associated with AD [7, 8], whereas physical exercise has been
proven to reduce both the progression of AD-like neuropathology in the TgCRND8
mouse model [9, 10] and the risk of AD [11], and improve cognitive function in AD
patients [12]. Many studies have been performed in both AD animal models and AD
patients to explore the molecular mechanisms underlying exercise-induced
improvement in memory or cognition. In several studies on AD animal models,
exercise induced significant reductions in A levels and tau
phosphorylation at multiple sites in both the cortex and hippocampus [13, 14, 15, 16, 17],
which was believed to be associated with inhibition of GSK3 activity via
enhancement in levels of p-GSK3/ ser21/9 [17, 18, 19]. However, in
a study on 6-month-old APP/PS1 transgenic (Tg) mice, housing mice in an enriched
environment for 4 weeks was unable to modulate GSK3 (ser9) activity in
either the hippocampus or the cortex [20]. The discrepancies in these results
might be due to variations in the forms of stimulation/exercise, intervention
times in the AD animals/patients (AD stage), and duration of the interventions.
In a previous study, 3-week voluntary wheel running in 15–19-month-old mice did
not significantly effect A levels [21]. However, in another study,
5-month treadmill exercise in 3- month and 12-month-old mice [15] significantly
reduced soluble A40/42 in mice of both ages and significantly reduced
A plaque burden in the 3-month-old mice but not the 12-month-old mice.
Therefore, intervention type and duration as well as AD stage seem to play
important roles in the outcomes.
To our knowledge, only a few studies have investigated the underlying molecular
mechanisms of memory function improvement in AD mouse models induced by aerobic
exercise with respect to regulation in protein kinase B (Akt)/GSK3 activity in both the cortex and
hippocampus. More importantly, no study has yet investigated the mechanisms in
term of GSK3/ activity in both the cortex and hippocampus of
moderate AD stage APP/PS1 mice. To fill these knowledge gaps, 6-month-old
APPSwe/PS1M146V double Tg AD mice and wild-type (Wt) mice were subjected to
12-week, moderate-intensity treadmill running. After the training, spatial
learning and memory were tested, and levels of A, phosphorylation of
tau, phosphorylation of Akt at ser473 (a major upstream modulator of GSK3), and
phosphorylation of GSK3 and GSK3 at tyr279/216 and ser21/9
were examined in tissues of both the cortex and hippocampus from both Tg mice and
Wt mice. By examining the effects of aerobic exercise on memory function in Tg
mice, and levels of A, phosphorylation of tau, phosphorylation of Akt at
ser473, and phosphorylation of GSK3 and GSK3 at tyr279/216 and
ser21/9 in both the cortex and hippocampus, we aimed to explore the molecular
mechanisms underlying the beneficial effects of physical exercise on AD
development.
2. Materials and Methods
2.1 Experimental Design
Male APPSwe/PS1M146V double-Tg mice and Wt mice were randomly assigned to a
sedentary group (Tg-Sed, Wt-Sed) or an aerobic exercise group (Tg-Ex, Wt-Ex).
After the physical training, all mice were subjected to an eight-arm radial maze
test for spatial learning and memory evaluation, then the mice were sacrificed
and brain tissues of the cortex and hippocampus were extracted for further
analysis.
2.2 Animals and Ethic Approval
Six-month-old male Tg mice (n = 20) and Wt male littermates (n = 20) were
purchased from Beijing HFK Bioscience Co., Ltd (Beijing, China). The mice were
kept under standard laboratory conditions (12:12 h light-dark cycle, at 22
2 °C and 45%–55% relative humidity) and provided with food
and water ad libitum. Both Tg mice and Wt mice were randomly and evenly
assigned to an exercise group (Tg-Ex, n = 10; Wt-Ex, n = 10) or a sedentary group
(Tg-Sed, n = 10; Wt-Sed, n = 10). Evidence has shown variations in
-amyloid pathological phenotype [22]; thus, to avoid sex-induced
variations in results, the study used only male APPSwe/PS1M146V double-Tg mice.
Ethics approval was obtained from the Ethics Committee of the Beijing Sport
University (2015015) and the Guiding Principles for Care and Use of Animals was
followed.
2.3 Aerobic Exercise Training
Before the 12-week exercise training program, both Wt-Ex and Tg-Ex mice were
familiarized with the treadmill training by running on a treadmill for 3
consecutive days at a speed of 10 m/min with a 0° inclination. After the
familiarization, each training session started with a standard warm-up by running
on the treadmill for 10 min at a speed of 12 m/min, followed by running for 50
min at a speed of 15 m/min, which was defined here as moderate intensity on the
basis of our previous studies where mice running at a speed of 12–15 meters/min
corresponded to an oxygen consumption of around 65–75% of maximal oxygen
consumption [23, 24, 25]. The training was carried out once per day, 5 days per week
for 12 consecutive weeks. Wt-Sed and Tg-Sed mice were placed on the treadmill for
10 min with the same time schedule as Wt-Ex and Tg-Ex mice but undertook no
exercise training.
2.4 Eight-Arm Radial Maze Test
On the next day following the last training session, all mice took an eight-arm
radial maze test in a quiet environment with weak light to facilitate the
assessment of spatial memory. The eight-arm radial maze apparatus (JLBehv-8ARMM,
Shanghai Jiliang, Shanghai, China) is comprised of eight arms, spaced
equidistantly, and visual reference cues were hung 1 m above the maze apparatus.
The test process has been previously described in detail [26]. Briefly, the test
includes 3 days of adaption and thereafter 10 consecutive days of testing. To
motivate the mice to seek the chocolate crumbs in the maze arms, the food supply
to the mice was reduced 3 days before the adaption so that the body weights of
the mice were reduced to 80–85% of their initial weights. Thereafter, the food
supply was maintained at such a level as to keep the body weights relatively
stable throughout the experiment [27, 28]. Body weights of the mice were measured
every other day to avoid 20% reduction. During the adaptation, the mice had 3
days to familiarize themselves with the test set by freely exploring the maze for
10 min with all eight of the arms baited with chocolate crumbs (0.08 g).
Before the test, chocolate crumbs were set in four randomly selected arms, and
the test began with putting the mice in the center of the platform heading
towards arm number one and ended when the mice had visited all the four baited
arms within a maximum time of 10 min. If the mice failed to visit all the baited
arms within 10 min, the mice would be excluded from group value calculations. The
test was conducted on each individual mouse for 10 consecutive days. Working
memory errors, reference memory errors, and time needed to complete each trial
were recorded. Evaluation of working memory error was performed by counting the
number of re-entries into a baited or non-baited arm, and similarly, evaluation
of reference memory error was performed by counting the number of entries into a
non-baited arm. For a specific mouse, the chocolate crumbs were placed in the
same four arms during the 10 trials. By the end of the tests, two mice in each
group failed to finish the test within the time limit, resulting in only eight
mice left in each group for statistical analysis.
2.5 Brain Tissue Processing and Immunostaining
The mice were anesthetized with isoflurane, then perfused with 4%
paraformaldehyde (P1110, Solarbio, Beijing, China) in 0.1M sodium phosphate
buffer (PBS) pH7.4 through the left ventricle of the heart. The brains were
removed from the skulls and postfixed by immersion in the same fixative for 48 h
at 4 °C. The cortical and hippocampal tissues were dissected and
dehydrated with 20% and 30% sucrose in 0.1 M PBS at 4 °C.
Subsequently, both tissues were frozen rapidly in liquid nitrogen and stored at
–80 °C until use. For immunostaining, cross sections (35
µm) were cut using a cryostat (CM1850, Leica, Wetzlar, Hessian,
Germany) and the sections were collected on glass slides. The sections from the
left hemisphere were used for immunohistochemistry, and the right hemisphere for
immunofluorescence. The immunofluorescence and immunohistochemistry staining
procedures have been previously described in detail [23, 29, 30]. The primary and
secondary staining antibodies are listed in Table 1.
Table 1.Antibodies used in Western Blotting, Immunohistochemistry and
Immunofluorescence staining.
| Antibody |
Primary antibody |
Secondary antibody |
Dilution |
Application |
| Antigen |
Dilution |
Host |
Isotype |
| ab131354 |
Tau phospho S262 |
1:5000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| ab109390 |
Tau phospho S396 |
1:5000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| ab32057 |
Tau |
1:5000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| ab28808 |
GSK3 phospho S21 |
1:5000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| ab131097 |
GSK3 phospho S9 |
1:10,000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| ab68476 |
GSK3(+) phospho (Y279+Y216) |
1:3000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| ab131344 |
GSK3 |
1:5000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| ab93926 |
GSK3 |
1:5000 |
Mouse |
IgG2a |
Goat Anti- Mouse IgG (H+L) |
1:20,000 |
Western Blot |
| ab81283 |
Akt phospho S473 |
1:10,000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| ab8805 |
Akt |
1:5000 |
Rabbit |
IgG |
Goat Anti- Rabbit IgG (H+L) |
1:20,000 |
Western Blot |
| 60004 -1-Ig |
GAPDH |
1:5000 |
Mouse |
IgG2b |
Goat Anti- Mouse IgG (H+L) |
1:20,000 |
Western Blot |
| SIG-39320 |
A |
1:500 |
Mouse |
IgG |
Goat anti-rabbit IgG-horseradish peroxidase |
1:500 |
Immunofluorescence |
| SIG-39320 |
A |
1:500 |
Mouse |
IgG |
Biotinylated goat anti-mouse IgG |
1:500 |
Immunohistochemistry |
Purchased from Abcam, Cambridge, UK; purchased from Proteintech
Group, Chicago, IL, USA; All the secondary antibodies were purchased from Jackson
Immuno Research Laboratories, West Grove, PA, USA. The primary and secondary
antibodies for western blotting were diluted in 5% bovine serum albumin in
Tris-buffered saline. GSK3, glycogen synthase kinase-3; A,
-amyloid; Akt, protein kinase B; GAPDH, glyceraldehyde-3-phosphate
dehydrogenase; IgG, immunoglobulin G.
The staining was examined under a microscope (IX71-F22PH, OLYMPUS, Tokyo, Japan)
connected to a computer equipped with a digital camera (modelDP71, OLYMPUS,
Tokyo, Japan). Image J software (National Institutes of Health, Bethesda, MD,
USA) was used for image analysis. A plaque levels were quantified on six
randomly selected areas of each section/mouse stained with immunofluorescence by
calculating the mean fluorescence intensity, which is the ratio of integrated
density to the area, using the same position on the section as a reference.
2.6 Western Blotting
The mice were anesthetized with isoflurane (26675-46-7, Sigma-Aldrich, Saint
Louis, MO, USA) then decapitated. After dissection of the cortex and hippocampus
on ice, the tissues were homogenized with phosphatase inhibitors and radio
immunoprecipitation assay lysis buffer (R0278, Sigma-Aldrich, St. Louis, MO,
USA). The protein extraction, protein content determination, and electrophoresis
procedures were the same as those described in our previous studies [23, 29, 30].
The primary and secondary antibodies are listed in Table 1.
Protein quantification was performed using Bio-Rad, ChemiDoc XRS+ System Image
Lab software (version 6.0.1, Bio-Rad, Hercules, CA, USA). The protein amounts were
expressed as semiquantitative ratios between the value of a specific protein and
the value of the reference protein glyceraldehyde-3-phosphate dehydrogenase
(GAPDH). Levels of microtubule-associated protein tau (p‐tau) ser262 and p‐tau ser396 were expressed as relative
values of total tau, levels of p-GSK3 ser21 and p-GSK3 tyr279
were expressed as relative values of total GSK3, and levels of
p-GSK3 ser9 and p-GSK3 tyr216 were expressed as relative values
of total GSK3.
2.7 Statistical Analysis
Statistical analysis was performed using SPSS software (version 22.0, SPSS Inc.,
Chicago, IL, USA). Group values of a specific variable were calculated based on
measurements of 10 consecutive days for all mice in the group. Repeated
measurement analysis of variance (ANOVA) was performed to analyze over-time
changes in the eight-arm radial maze test performance among groups (i.e., Wt-Sed,
Wt-Ex, Tg-Sed, and Tg-Ex), followed by Bonferroni post-hoc analysis when group,
time, and group-time interaction were significant. Two-way ANOVA (with genotype
[Wt or Tg] and exercise [Sed or Ex] as two factors) was used to analyze the
results of western blotting, and a simple main effect (multiple comparison
adjusted by the Bonferroni approach) was reported when genotype-exercise
interaction was significant. The student’s t-test was applied for the
analysis of immunofluorescence staining between Tg-Sed and Tg-Ex mice. A
probability of less than 0.05 was used to reject the null hypothesis and deemed
to indicate a statistically significant difference. All data are expressed as
mean (standard deviation) (mean (SD)), except for the A plaque level
data, which was expressed as mean standard error of the mean (mean
SEM).
3. Results
3.1 Eight-Arm Radial Maze Test
Repeated-measures ANOVA revealed that test days and groups had significant
effects on the working memory errors (Fig. 1A) and reference memory errors (Fig. 1B). Both working memory errors (test days: F = 6.317, p
0.001) and reference memory errors (test days: F = 2.144, p
0.001) were significantly decreased following the progress of the test in all the
four groups, and significantly different between groups (groups: working memory
errors, F = 8.783, p 0.001; reference memory errors,
F = 13.543, p 0.001). Interaction between groups and test
days had no significant effect on either working memory errors or reference
memory errors (groups test days: working memory errors, F =
1.095, p = 0.361; reference memory errors, F = 0.489,
p = 0.957). The main effect of test days and groups revealed that Tg-Sed
mice showed significantly higher working memory errors (p 0.001) and
reference memory errors (p = 0.003) than Wt-Sed mice, and Tg-Ex mice
showed significantly lower working memory errors (p = 0.018) and
reference memory errors (p = 0.042) than Tg-Sed mice (Fig. 1A,B).
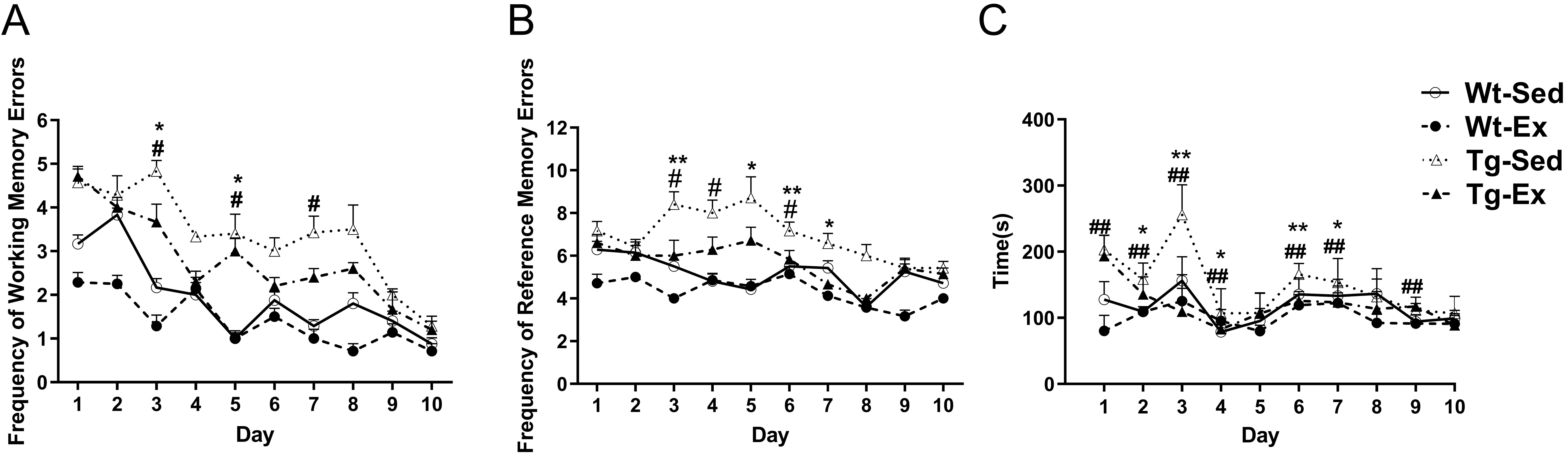 Fig. 1.
Fig. 1.
Results of the eight-arm radial maze test. Both
working memory errors (A) and reference memory errors (B) were significantly
decreased following the progress of the test in all four groups (n = 10/group;
test days: working memory errors, F = 6.317, p 0.001;
reference memory errors, F = 2.144, p 0.001) and were
significantly different between groups (groups: working memory errors, F
= 8.783, p 0.001; reference memory errors, F = 13.543,
p 0.001). Tg-Sed mice (n = 8) showed significantly higher working
memory errors and reference memory errors than Wt-Sed mice (n = 8) on three test
occasions. Tg-Ex mice (n = 8) had significantly lower working memory errors on
two test occasions, and reference memory errors than Tg-Sed mice (n = 8) on four
test occasions. Time needed to complete the test was significantly decreased
following the progress of the test for all four groups (n = 10/group; test days:
F = 64.724, p 0.001) and significantly different between
groups (C; groups: F = 103.1, p 0.001). Tg-Sed mice needed
significantly longer time to complete the test than Wt-Sed mice on seven test
occasions, whereas Tg-Ex mice needed significantly shorter time to complete the
test than Tg-Sed mice on five test occasions (C). Data are expressed as mean
(SD). Note: Wt, wild-type; Tg, transgenic; Sed, sedentary; Ex, exercise; SD,
standard deviation. p 0.05, Tg-Ex vs Tg-Sed; p 0.01, Tg-Ex vs Tg-Sed; p 0.05, Tg-Sed vs Wt-Sed;
p 0.01, Tg-Sed vs Wt-Sed.
The time needed to complete each repetition of the test is shown in Fig. 1C. The
time decreased significantly following the progress of the test for all four
groups (test days: F = 64.724, p 0.001), and was
significantly different between groups (groups: F = 103.1,
p 0.001). Significant interaction between groups and test days was
also found (groups test days: F = 14.746, p 0.001).
The Bonferroni post-hoc analysis revealed that Tg-Sed mice needed significantly
longer time to complete the test on day 1 (p 0.01), day 2
(p 0.01), day 3 (p 0.01), day 4 (p 0.01),
day 6 (p 0.01), day 7 (p 0.01), and day 9 (p 0.01) than Wt-Sed mice on the corresponding days. In contrast, Tg-Ex mice
needed significantly shorter time to complete the test on day 2 (p
0.05), day 3 (p 0.01), day 4 (p 0.05), day 6
(p 0.01), and day 7 (p 0.05) than Tg-Sed mice on the
corresponding days.
3.2 A Plaques
A plaques were not observed in either the cortex or hippocampus of
Wt-Sed or Wt-Ex mice, but were in both tissues of both Tg-Sed and Tg-Ex mice
(Fig. 2A,B). Statistical analysis revealed significantly lower levels of
A plaques in both tissues of Tg-Ex mice than in respective tissues of
Tg-Sed mice (Fig. 2C).
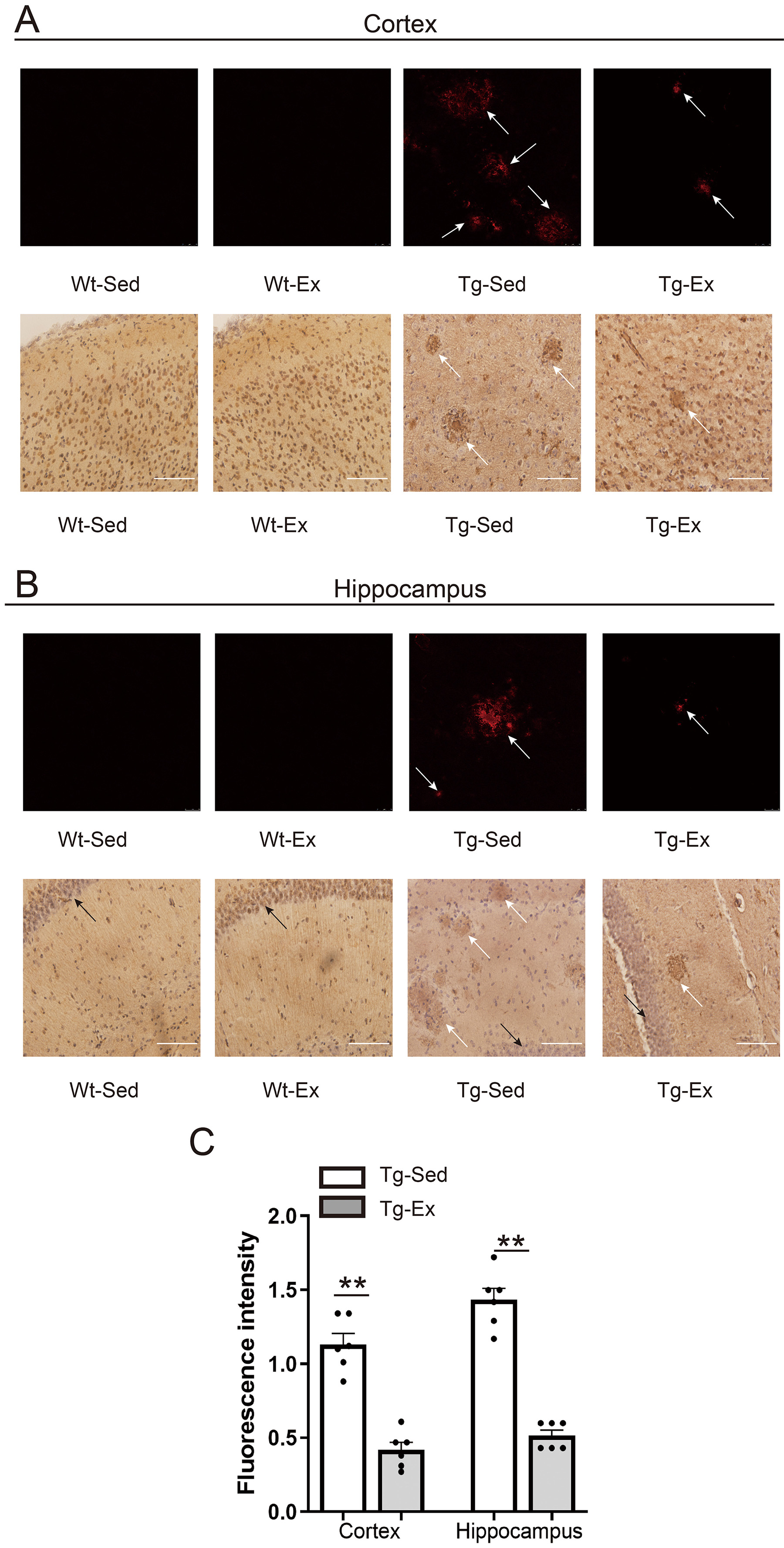 Fig. 2.
Fig. 2.
A plaque levels in the cortex and hippocampus.
Representative images of immunofluorescence staining (upper panel) and
immunohistochemical staining (lower panel) in the cortex (A) and hippocampus (B),
revealing A plaques (white arrows) in the cortex and hippocampus, and
pyramidal layer in the hippocampus (black arrows). Quantification of the
fluorescence staining for A plaques and comparisons of the results
between Tg-Sed and Tg-Ex mice in respective tissues of the cortex and hippocampus
are shown in the histogram (C). In both the cortex and hippocampus, Tg-Ex mice
had significantly lower levels of A plaques than in the respective
tissues of Tg-Sed mice (n = 6/group). Data are expressed as mean SEM.
Note: Wt, wild-type; Tg, transgenic; Sed, sedentary; Ex, exercise; SEM, standard
error of the mean.p 0.01 (Tg-Ex vs Tg-Sed). Scale bar = 25
µm.
3.3 Tau Phosphorylation
Western blot analysis revealed four different protein bands with different
molecular weights, representing phosphorylation of total tau, p-tau ser262, p-tau
ser396, and reference protein GAPDH, respectively, in the cortex (Fig. 3 upper
panel, left) and hippocampus (Fig. 3 upper panel, right; all original figures of
Western Blot can be found in the Supplementary Material) of Wt-Sed,
Wt-Ex, Tg-Sed, and Tg-Ex mice.
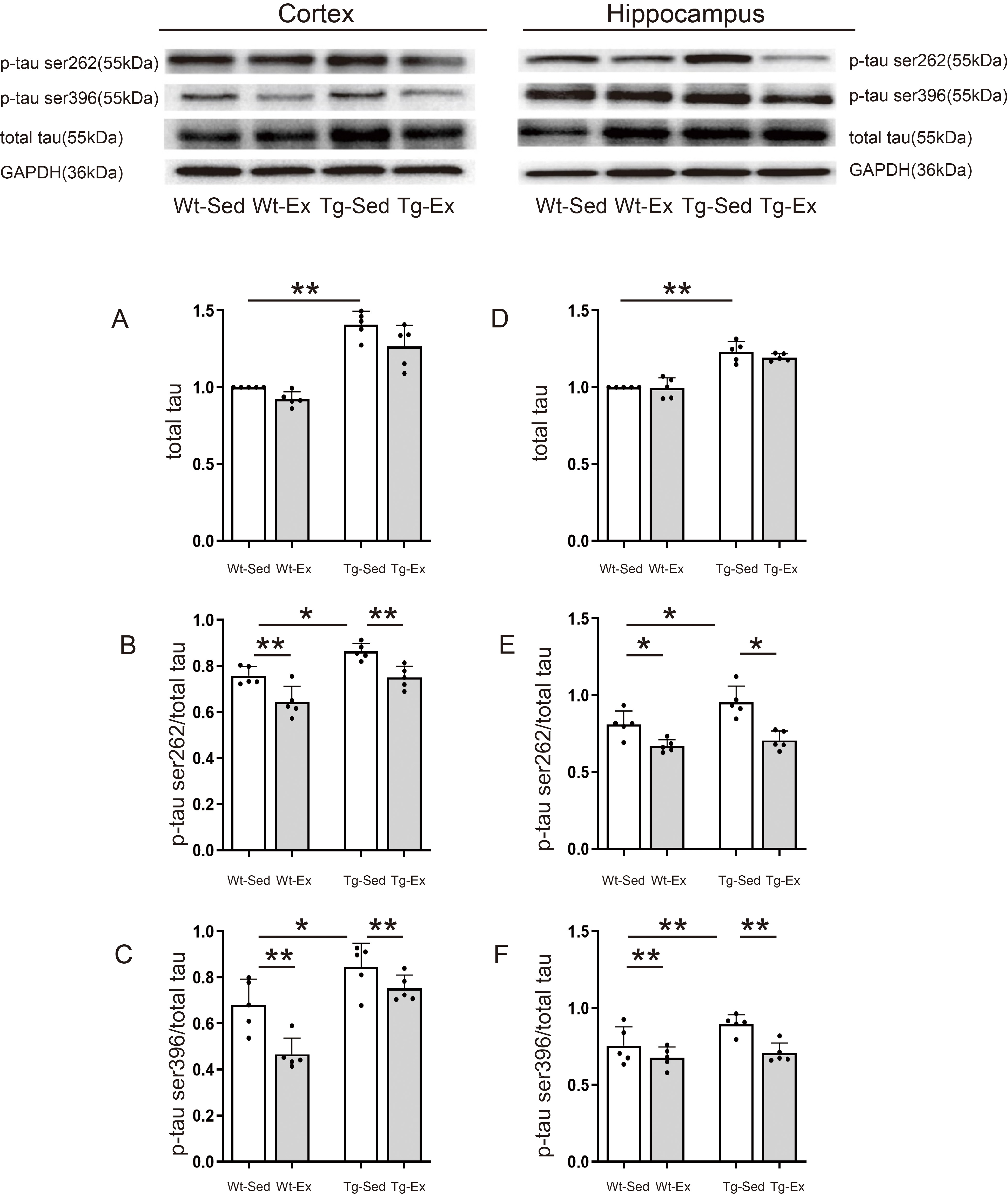 Fig. 3.
Fig. 3.
Tau phosphorylation levels in the cortex and hippocampus.
Representative images of western blots for tau (A,D), p-tau ser262 (B,E), and
p-tau ser396 (C,F) in the cortex (upper panel, left) and hippocampus (upper panel, right) (n = 5/group). Tau in the cortex
(A) and hippocampus (D) was significantly increased in Tg-Sed mice compared with
Wt-Sed mice and this increase was not blocked by the treadmill exercise. P-tau
ser262/tau (B) and p-tau ser396/tau (C) in the cortex were significantly
increased in Tg-Sed mice compared with Wt-Sed mice but significantly decreased in
Tg-Ex mice compared with Tg-Sed mice and Wt-Ex mice compared with Wt-Sed mice.
P-tau ser262/tau in the hippocampus (E) was significantly increased in Tg-Sed
mice compared with Wt-Sed mice but significantly decreased in Tg-Ex mice compared
with Tg-Sed mice and Wt-Ex mice compared with Wt-Sed mice. P-tau ser396/tau in
the hippocampus (F) was significantly increased in Tg-Sed mice compared with
Wt-Sed mice but significantly decreased in Tg-Ex mice compared with Tg-Sed mice
and Wt-Ex mice compared with Wt-Sed mice. Data are expressed as mean (SD). Note:
Wt, wild-type; Tg, transgenic; Sed, sedentary; Ex, exercise; SD, standard
deviation. *p 0.05; **p 0.01.
Tau phosphorylation levels and their comparisons between different groups are
shown in Fig. 3A–F.
Two-way ANOVA revealed that genotype had significant effects on total tau in
both the cortex (genotype: F = 72.203, p 0.001) and hippocampus
(genotype: F = 98.128, p 0.001), but there were no significant
exercise effects or interaction between genotype and exercise on total tau in
both the cortex (exercise: F = 5.012, p = 0.061; genotype
exercise: F = 0.007, p = 0.935) and hippocampus
(exercise: F = 0.885, p = 0.361; genotype exercise:
F = 0.572, p = 0.460). The main effect of genotype revealed
that Tg groups had significantly higher levels of total tau in both the cortex
and hippocampus than in respective tissues of Wt groups (Fig. 3A,D).
Two-way ANOVA revealed that genotype and exercise had significant effects on the
levels of p-tau ser262 in both the cortex (genotype: F = 8.348,
p = 0.012; exercise: F = 8.820, p = 0.009) and
hippocampus (genotype: F = 6.981, p = 0.021; exercise:
F = 7.098, p = 0.017), but there was no significant interaction
between genotype and exercise on the levels of p-tau ser262 in both the cortex
(genotype exercise: F = 0.028, p = 0.870) and
hippocampus (genotype exercise: F = 3.967, p =
0.070). The main effect of genotype and exercise revealed that Tg groups had
significantly higher levels of p-tau ser262 in both the cortex and hippocampus
than in respective tissues of Wt groups. Exercise groups had significantly lower
levels of p-tau ser262 in both the cortex and hippocampus than in respective
tissues of sedentary groups (Fig. 3B,E).
Two-way ANOVA revealed that genotype and exercise had significant effects on the
levels of p-tau ser396 in both the cortex (genotype: F = 5.821,
p = 0.031; exercise: F = 19.320, p = 0.001) and
hippocampus (genotype: F = 46.984, p = 0.001; exercise:
F = 15.278, p = 0.002), but there was no significant
interaction between genotype and exercise on the levels of p-tau ser396 in both
the cortex (genotype exercise: F = 0.981, p = 0.335)
and hippocampus (genotype exercise: F = 3.078, p =
0.103). The main effect of genotype and exercise revealed that Tg groups had
significantly higher levels of p-tau ser396 in both the cortex and hippocampus
than in respective tissues of Wt groups. Exercise groups had significantly lower
levels of p-tau ser396 in both the cortex and hippocampus than in respective
tissues of sedentary groups (Fig. 3C,F).
3.4 GSK3 Kinase Activity in the Cortex and Hippocampus
Western blotting revealed three different protein bands with different molecular
weights, representing total GSK3, p-GSK3
ser21/p-GSK3 tyr279, and reference protein GAPDH, respectively, in both
tissues of each group (Fig. 4 upper panel). Quantification of the bands and
comparisons of both p-GSK3 ser21 and p-GSK3 tyr279 levels in
both tissues between different groups are shown in the histogram (Fig. 4A–D).
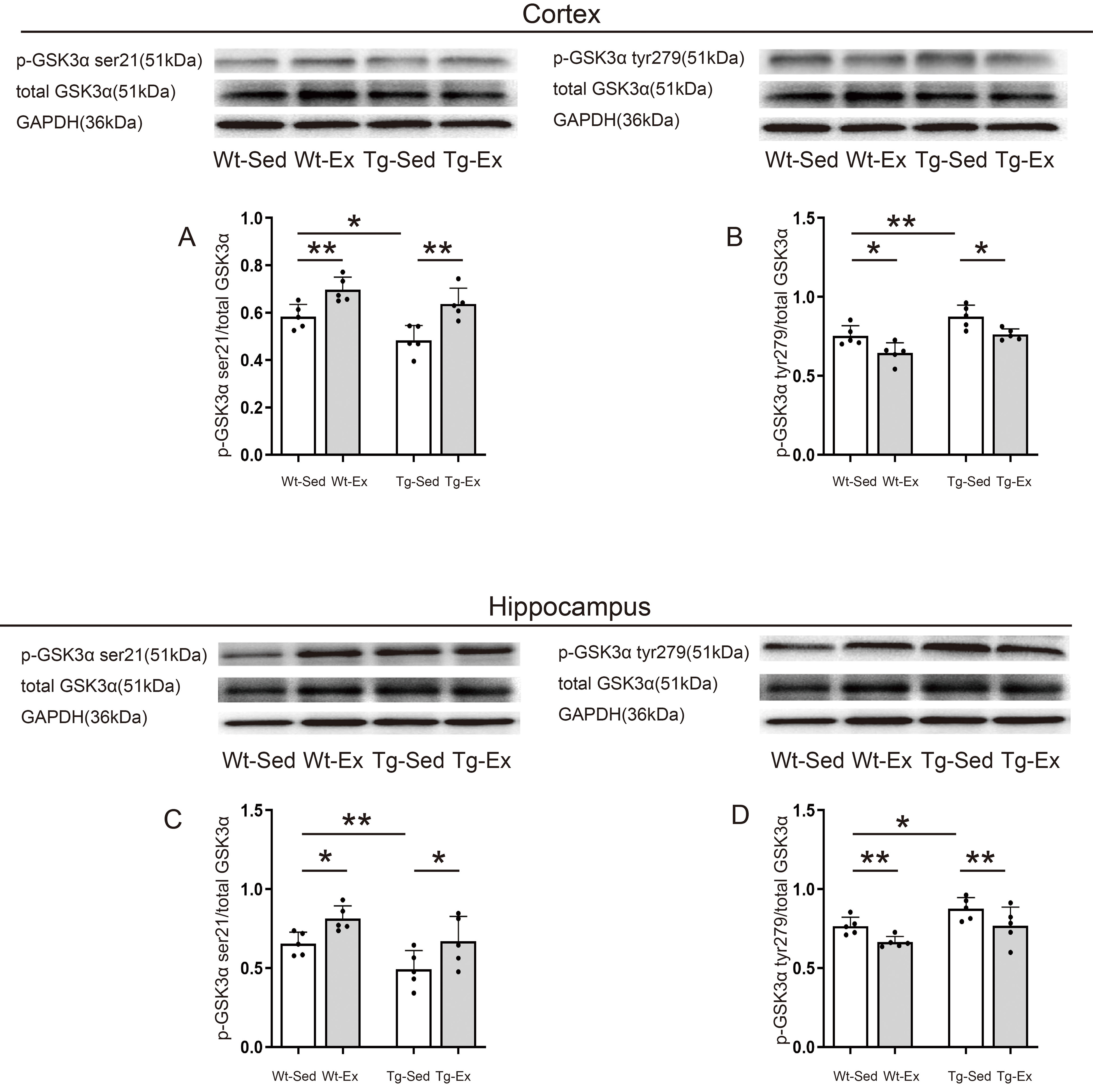 Fig. 4.
Fig. 4.
GSK3 kinase activity in the cortex and hippocampus.
Representative images of western blots for p-GSK3 ser21 (A,C),
p-GSK3 tyr279 (B,D), and GSK3 (A–D) in the cortex and
hippocampus (n = 5/group). In the cortex of Tg-Sed mice, p-GSK3
ser21/GSK3 was significantly decreased compared with that of Wt-Sed
mice but significantly increased in Tg-Ex mice compared with Tg-Sed mice and
Wt-Ex mice compared with Wt-Sed mice (A). P-GSK3 tyr279/GSK3
in the cortex was significantly increased in Tg-Sed compared with Wt-Sed but
significantly decreased in Tg-Ex compared with Tg-Sed mice and Wt-Ex mice
compared with Wt-Sed mice (B). P-GSK3 ser21/GSK3 in the
hippocampus was significantly decreased in Tg-Sed mice compared with Wt-Sed mice
but significantly increased in Tg-Ex mice compared with Tg-Sed mice and Wt-Ex
mice compared with Wt-Sed mice (C). P-GSK3 tyr279/GSK3 in the
hippocampus was significantly increased in Tg-Sed mice compared with Wt-Sed mice
but significantly decreased in Tg-Ex mice compared with Tg-Sed mice and Wt-Ex
mice compared with Wt-Sed mice (D). Data are expressed as mean (SD). Note: Wt,
wild-type; Tg, transgenic; Sed, sedentary; Ex, exercise; SD, standard deviation.
*p 0.05; **p 0.01.
Two-way ANOVA revealed that genotype and exercise had significant effects on the
levels of p-GSK3 ser21 (genotype: F = 7.654, p =
0.017; exercise: F = 12.397, p = 0.004) and p-GSK3
tyr279 (genotype: F = 17.736, p = 0.001; exercise: F =
8.980, p = 0.011) in the cortex, but there was no significant
interaction between genotype and exercise on the levels of p-GSK3 ser21
(genotype exercise: F = 0.713, p = 0.415) and
p-GSK3 tyr279 (genotype exercise: F = 0.003,
p = 0.955) in the cortex. The main effect of genotype and exercise
revealed that Tg groups had significantly lower levels of p-GSK3 ser21
and higher levels of p-GSK3 tyr279 in the cortex than in respective
tissues of Wt groups. Exercise groups had significantly higher levels of
p-GSK3 ser21 and lower levels of p-GSK3 tyr279 in the cortex
than in respective tissues of sedentary groups (Fig. 4A,B).
Two-way ANOVA revealed that genotype and exercise had significant effects on the
levels of p-GSK3 ser21 (genotype: F = 9.573, p =
0.009; exercise: F = 6.217, p = 0.028) and p-GSK3
tyr279 (genotype: F = 5.397, p = 0.035; exercise: F =
33.209, p 0.001) in the hippocampus, but there was no significant
interaction between genotype and exercise on the levels of p-GSK3 ser21
(genotype exercise: F = 0.684, p = 0.424) and
p-GSK3 tyr279 (genotype exercise: F = 0.156,
p = 0.699) in the hippocampus. The main effect of genotype and exercise
revealed that Tg groups had significantly lower levels of p-GSK3 ser21
and higher levels of p-GSK3 tyr279 in the hippocampus than in
respective tissues of Wt groups. Exercise groups had significantly higher levels
of p-GSK3 ser21 and lower levels of p-GSK3 tyr279 in the
hippocampus than in respective tissues of sedentary groups (Fig. 4C,D).
P-GSK3 ser9 and p-GSK3 tyr216 levels in both tissues are shown
in Fig. 5. Western blotting revealed three different protein bands with different
molecular weights, representing total GSK3, p-GSK3ser9/p-GSK3 tyr216, and reference protein GAPDH, respectively, in both
tissues of each individual group (upper panel in Fig. 5A–D). Quantification of
the bands and comparisons between different groups are shown in the histograms
(Fig. 5A–D).
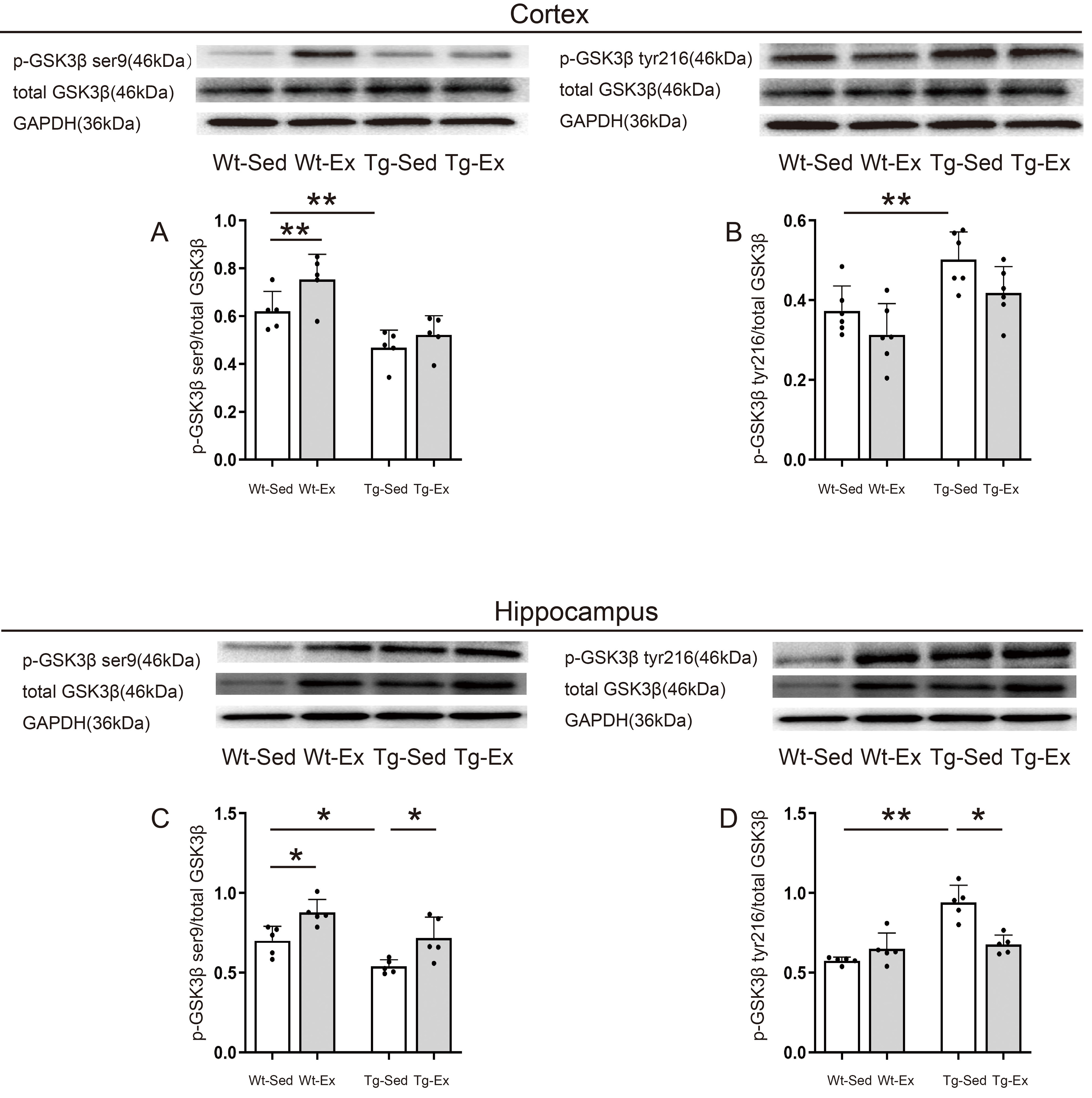 Fig. 5.
Fig. 5.
GSK3 kinase activity in the cortex and hippocampus.
Representative images of western blots for p-GSK3 ser9 (A,C),
p-GSK3 tyr216 (B,D), and GSK3 (A–D) in the cortex and
hippocampus (n = 5/group). P-GSK3 ser9/GSK3 in the cortex was
significantly decreased in Tg-Sed mice compared with Wt-Sed mice but only
increased in Wt-Ex mice compared with Wt-Sed (A). P-GSK3
tyr216/GSK3 in the cortex was significantly increased in Tg-Sed mice
compared with Wt-Sed mice but similar between Tg-Ex and Tg-Sed mice, and between
Wt-Ex and Wt-Sed mice (B). P-GSK3 ser9/GSK3 in the hippocampus
was significantly decreased in Tg-Sed mice compared with Wt-Sed mice but
significantly increased in Tg-Ex mice compared with Tg-Sed mice and Wt-Ex mice
compared with Wt-Sed mice (C). P-GSK3 tyr216/GSK3 in the
hippocampus was significantly increased in Tg-Sed mice compared with Wt-Sed mice
but significantly decreased in Tg-Ex mice compared with Tg-Sed mice, and similar
between Wt-Ex and Wt-Sed mice (D). Data are expressed as mean (SD). Note: Wt,
wild-type; Tg, transgenic; Sed, sedentary; Ex, exercise; SD, standard deviation.
*p 0.05; **p 0.01.
Two-way ANOVA revealed that genotype had significant effects on the levels of
p-GSK3 ser9 (genotype: F = 34.192, p = 0.001) and
p-GSK3 tyr216 (genotype: F = 15.190, p = 0.001) in the
cortex. Significant exercise effects and interaction between genotype and
exercise were observed on the levels of p-GSK3 ser9 (exercise:
F = 11.851, p = 0.003; genotype exercise: F
= 5.349, p = 0.034) but not on p-GSK3 tyr216 (exercise:
F = 3.952, p = 0.064; genotype exercise: F
= 0.095, p = 0.762) in the cortex. The main effect of genotype revealed
that Tg groups had significantly lower levels of p-GSK3 ser9 and higher
levels of p-GSK3 tyr216 in the cortex than in respective tissues of Wt
groups. The multiple comparison test revealed that Wt-Ex mice had significantly
higher levels of p-GSK3 ser9 (p 0.01) than Wt-Sed mice
(Fig. 5A).
Two-way ANOVA revealed that genotype and exercise had significant effects on the
levels of p-GSK3 ser9 (genotype: F = 5.821, p = 0.034;
exercise: F = 6.871, p = 0.024) and p-GSK3 tyr216
(genotype: F = 39.914, p = 0.001; exercise: F = 8.074,
p = 0.019; genotype exercise: F = 10.467, p
= 0.010) in the hippocampus, but there was no significant interaction between
genotype and exercise on the levels of p-GSK3 ser9 (genotype
exercise: F = 0.393, p = 0.637) in the hippocampus. The main
effect of genotype and exercise revealed that Tg groups had significantly lower
levels of p-GSK3 ser9 in the hippocampus than in respective tissues of
Wt groups. Exercise groups had significantly higher levels of p-GSK3
ser9 in the hippocampus than in respective tissues of sedentary groups (Fig. 5C).
The multiple comparison test revealed that Tg-Sed mice had significantly higher
levels of p-GSK3 tyr216 (p 0.01) than Wt-Sed mice (Fig. 5D). In contrast, Tg-Ex mice (p = 0.011) had significantly lower levels
of p-GSK3 tyr216 than Tg-Sed mice, and similar levels of p-GSK3
tyr216 in Wt-Ex and Wt-Sed mice (p = 0.785) (Fig. 5D).
3.5 Akt Phosphorylation in the Cortex and Hippocampus
Western blotting revealed three different protein bands with different molecular
weights, representing p-Akt ser473, total Akt, and reference protein GAPDH,
respectively, in both tissues of each group (Fig. 6 upper panel). Quantification
of the bands and comparisons between different groups are shown in Fig. 6.
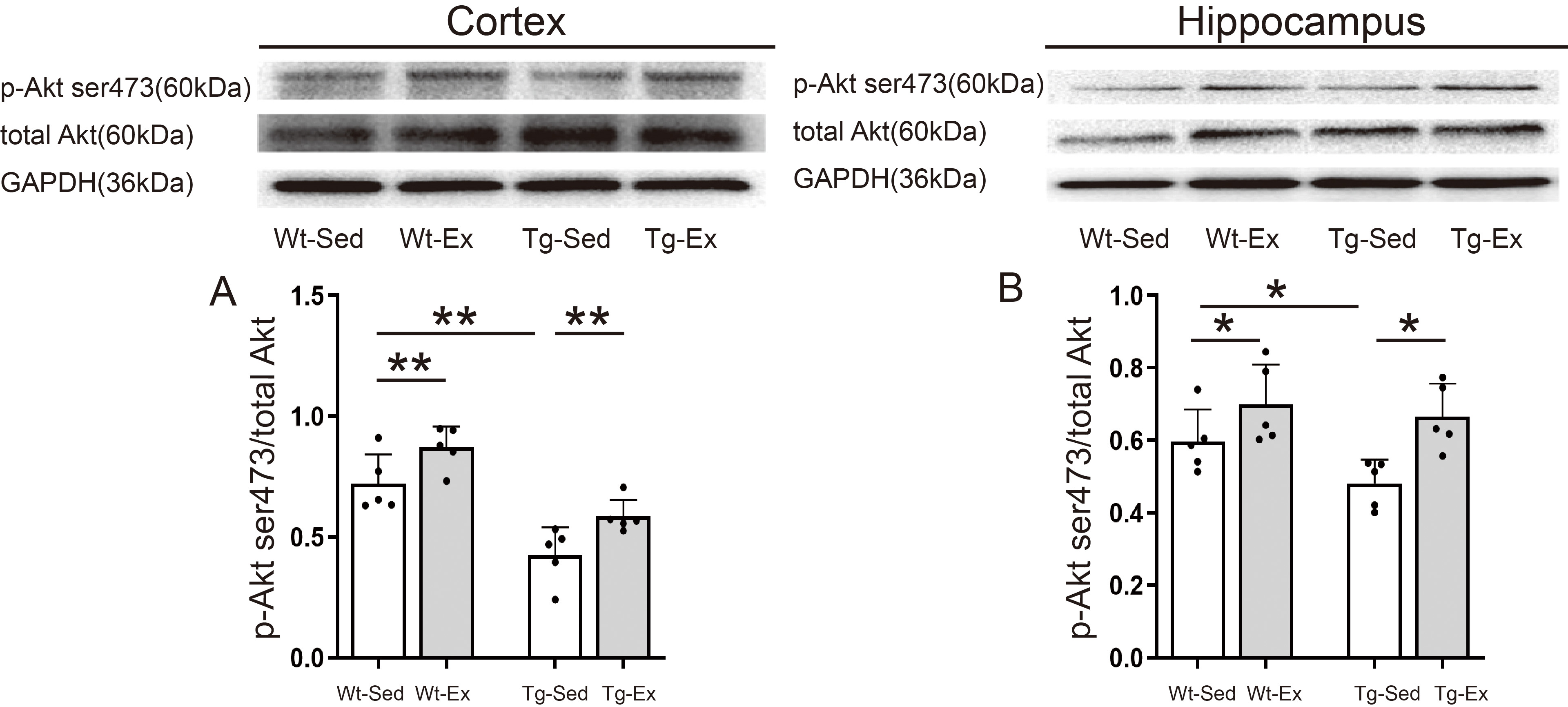 Fig. 6.
Fig. 6.
Akt kinase activity in the cortex and hippocampus.
Representative images of western blots of p-Akt ser473, total Akt, and reference
protein GAPDH in the cortex (upper panel, left) and hippocampus (upper panel,
right; n = 5/group). P-Akt ser473/Akt in the cortex was significantly decreased
in Tg-Sed mice compared with Wt-Sed mice but significantly increased in Tg-Ex
mice compared with Tg-Sed mice and Wt-Ex mice compared with Wt-Sed mice (A).
P-Akt ser473/Akt in the hippocampus was significantly decreased in Tg-Sed mice
compared with Wt-Sed mice, and both significantly increased in Tg-Ex and Wt-Ex
mice (B). Data are expressed as mean (SD). Note: Wt, wild-type; Tg, transgenic;
Sed, sedentary; Ex, exercise; SD, standard deviation. *p 0.05;
**p 0.01.
Two-way ANOVA revealed that genotype and exercise had significant effects on the
levels of p-Akt ser473 in the cortex (genotype: F = 38.969, p =
0.001; exercise: F = 12.950, p = 0.004) and hippocampus
(genotype: F = 5.484, p = 0.044; exercise: F = 5.202,
p = 0.048), but there was no significant interaction between genotype
and exercise on the levels of p-Akt ser473 in the cortex (genotype
exercise: F = 0.221, p = 0.648) and hippocampus (genotype
exercise: F = 1.433, p = 0.262). The main effect of
genotype and exercise revealed that Tg groups had significantly lower levels of
p-Akt ser473 in both the cortex and hippocampus than in respective tissues of Wt
groups. Exercise groups had significantly higher levels of p-Akt ser473 in both
the cortex and hippocampus than in respective tissues of sedentary groups (Fig. 6A,B).
4. Discussion
The study revealed that moderate-stage AD APP/PS1 mice had significantly
impaired memory function, along with significantly higher levels of A
plaques, total tau, p-tau ser262, and p-tau ser396 in both the cortex and
hippocampus than Wt mice, whereas 12-week, moderate-intensity treadmill exercise
in AD mice induced significant improvements in memory function together with a
significant reduction in A plaques and all the three tau molecules in
both tissues. Importantly, the moderate-stage AD APP/PS1 mice had significantly
lower levels of both p-GSK3 ser21 and p-GSK3 ser9, but higher
levels of both p-GSK3 tyr279 and p-GSK3 tyr216 in both tissues
than Wt sedentary mice. However, after the 12-week exercise training, the
hippocampus had significantly higher levels of both p-GSK3 ser21 and
p-GSK3 ser9 but lower levels of both p-GSK3 tyr279 and
p-GSK3 tyr216, whereas the cortex had significantly higher levels of
p-GSK3 ser21 but only lower levels of p-GSK3 tyr279, in
comparison with sedentary AD mice. The results strongly suggest the effectiveness
of the training in improving memory function in the AD mice, which was believe
occurred most likely through differential regulation of the Akt/GSK3 signaling
pathways in the cortex and hippocampus.
The double-Tg APPSwe/PS1M146V mouse strain is currently widely used in studying
the pathogenesis of AD, and impaired memory in these mice has been reported
previously [31, 32, 33]. In our study, we observed significantly impaired memory
function in the moderate-stage AD APP/PS1 mice. However, the impaired memory was
rescued mice after the 12-week aerobic exercise training program. This is
consistent with previous studies where regular exercise led to cognitive
improvement/reservation, as reflected by lower working memory errors and
reference memory errors [34, 35], and shorter time to complete the Maze test in
other AD mouse models such as the triple-Tg and TgCRND8 lines. Regular exercise
also decreased completion time of the Floor Maze test in AD patients [36, 37].
However, contradictory results showing no improvement in cognitive performance
after regular exercise have also been observed in 10-month-old APP/PS1 mice,
9-month-old triple-Tg AD female mice, and AD patients [38, 39, 40, 41]. The reasons for
this are attributed to many different factors such as exercise type and
intensity, health status, and wide range of Tg animal strains and AD stages
[34, 36, 37, 42]. The present observation of significant improvement in memory
function in the moderate-stage AD stage Tg-Ex mice suggests both the efficiency
of the training protocol and the increased susceptibility of the mice to the
training, i.e., memory function can be improved in moderate-stage AD APP/PS1
mice.
The double-Tg APPSwe/PS1M146V mouse strain is characterized by initial and
progressive enhancement of A plaques [31, 32, 33]. Indeed, clusters of
A plaques were clearly observed in both the cortex and hippocampus of
the 6-month-old AD mice, similar to the observations in double-Tg APPSwe/PS1M146V
mice [31, 32, 33]. However, the 12-week moderate-intensity exercise training program
induced a significant reduction in A plaques in both tissues, suggesting
that moderate-intensity aerobic exercise is effective in regulating A
plaque production in moderate-stage AD mice. Similar results have been observed
in other AD mouse models where exercise training in Tg2576 AD mice resulted in
lower A plaques in both the cortex and hippocampus [43], and enriched
housing of TgCRND8 AD mice induced the reduction of A plaque burden
[44].
Intracellular accumulation of phosphorylated tau is another hallmark lesion in
AD [45]. The present study extensively studied tau by examining total tau, p-tau
ser262, and p-tau ser396 in both the cortex and hippocampus. The results revealed
that while the AD mice had significantly higher levels of total of tau, p-tau
ser262, and p-tau ser396 in both the cortex and hippocampus than in that of Wt
mice, moderate-intensity exercise training induced significant reductions in all
three tau molecules in both tissues (see Fig. 3). The results are consistent with
the observations on A plaques in AD-exercised mice, confirming the
effectiveness of the exercise in preventing or delaying deterioration of AD
development in moderate-stage AD mice. Interestingly, the study revealed that the
exercise also induced a significant reduction in p-tau ser262 and p-tau ser396 in
both the cortex and hippocampus of Wt mice compared with Wt sedentary mice,
indicating the significance of physical exercise in preventing the occurrence of
AD. Similar to the present observations, exercise-induced reduction in A
plaques and tau phosphorylation have been observed previously [13, 16], along with
improvement in cognitive function and behaviors [16, 17, 46]. Considering the
observations in the improvement in memory function, and the reductions in
A plaques and tau molecules in the AD-exercised mice, we assume that the
reductions are closely associated with the improvement in memory function.
Nonetheless, it is worth noting that a previous study on another AD mouse model
revealed that exercise had no significant effect on 12-month TgCRND8 mice in
abnormal APP metabolism, tau pathology, or angiogenesis [12].
To explore the molecular mechanisms underlying the improvement in memory
function and the significant reduction in A plaques in both tissues of
AD mice following a 12-week exercise training program, the study further examined
Akt/GSK3 signaling pathways. GSK3 deregulation has been proven to account for
memory impairment, increased A production, hyper-phosphorylation of tau,
and inflammatory responses in AD [47]. Indeed, our study revealed that the AD
sedentary mice had significantly lower levels of both p-GSK3 ser21 and
p-GSK3 ser9, but higher levels of both p-GSK3 tyr279 and
p-GSK3 tyr216 (see Figs. 4,5) in both tissues, along with significantly
lower memory function than Wt sedentary mice. As p-GSK3 tyr279 and
p-GSK3 tyr216 function to activate GSK3, whereas p-GSK3 ser21
and p-GSK3 ser9 inhibit GSK3 [48], the reduced levels of
p-GSK3 ser21 and p-GSK3 ser9 together with the enhanced levels
of p-GSK3 tyr279 and p-GSK3 tyr216 in the AD sedentary mice
are believed to work coordinately to activate GSK3 and GSK3,
leading to A plaque formation and resulting in enhanced A
plaque levels (see Fig. 3). These findings suggest that GSK3 and
GSK3 phosphorylation were similar in both tissues, although A
pathology occurred earlier in the cortex than the hippocampus.
After the 12-week exercise training program in the AD mice, the hippocampus had
significantly higher levels of both p-GSK3 ser21 and p-GSK3
ser9 but lower levels of both p-GSK3 tyr279 and p-GSK3 tyr216,
whereas the cortex had significantly higher levels of p-GSK3 ser21 but
lower levels of p-GSK3 tyr279 in comparison to respective tissues of AD
sedentary mice (see Figs. 4,5). The activity of GSK3/ is
believed to be maintained through phosphorylation at both tyr279/216 sites
(activation) and ser21/9 sites (inhibition) [49], though the activity of GSK3 has
also been suggested to be regulated through phosphorylation at Ser9/21, rather
than Tyr 216/279 [50]. Thus, in both tissues of the exercised-AD mice, the
enhanced levels of both p-GSK3 ser21 and p-GSK3 ser9 together
with the reduced levels of both p-GSK3 tyr279 and p-GSK3
tyr216 are believed to work coordinately to result in the reduction in GSK3
activity, which subsequently led to the decrease in A plaques and tau
phosphorylation. However, the exercise training had significant impact on both
GSK3 and GSK3 in the hippocampus, but not on GSK3 in the cortex of the AD mice, indicating that
the aerobic exercise is sufficient to suppress the GSK3 signal in the
hippocampus, but not the cortex of moderate-stage AD mice.
To further explore the molecular signaling pathways of GSK3 activation, we
examined the activity of the GSK3 upstream regulator Akt. The results revealed
that both tissues of AD sedentary mice had significantly lower p-Akt ser473
levels than respective tissues of Wt sedentary mice (see Fig. 6). However, after
the 12-week exercise training program, both tissues of the AD mice showed
significantly higher levels of p-Akt ser473 than respective tissues of AD
sedentary mice. As GSK3 could be inactivated by Akt ser473 phosphorylation
[51, 52], the increased p-Akt ser473 levels indicated enhanced inhibition of GSK3
activity in both tissues of AD exercised-mice.
Considering all the results, we propose a scenario of molecular reactions in the
two regions of the brain of the APP/PS1 mice in response to the exercise training
as shown in Fig. 7: the exercise induced enhancement in p-Akt ser473 in both the
cortex and hippocampus, which subsequently led to enhancement in inhibition of
both GSK3 and GSK3 by increasing both p-GSK3 ser21
and p-GSK3 ser9 but decreasing both p-GSK3 tyr279 and
p-GSK3 tyr216 in the hippocampus, as well as by increasing
p-GSK3 ser21 but decreasing p-GSK3 tyr279 in the cortex. The inhibition of GSK3 and GSK3 activity
in the cortex and hippocampus resulted in the reduction in both p-tau ser396 and
p-tau ser262, as well as in A plaques in both regions, and eventually
improved memory function or delayed memory deterioration in the AD-exercised
mice.
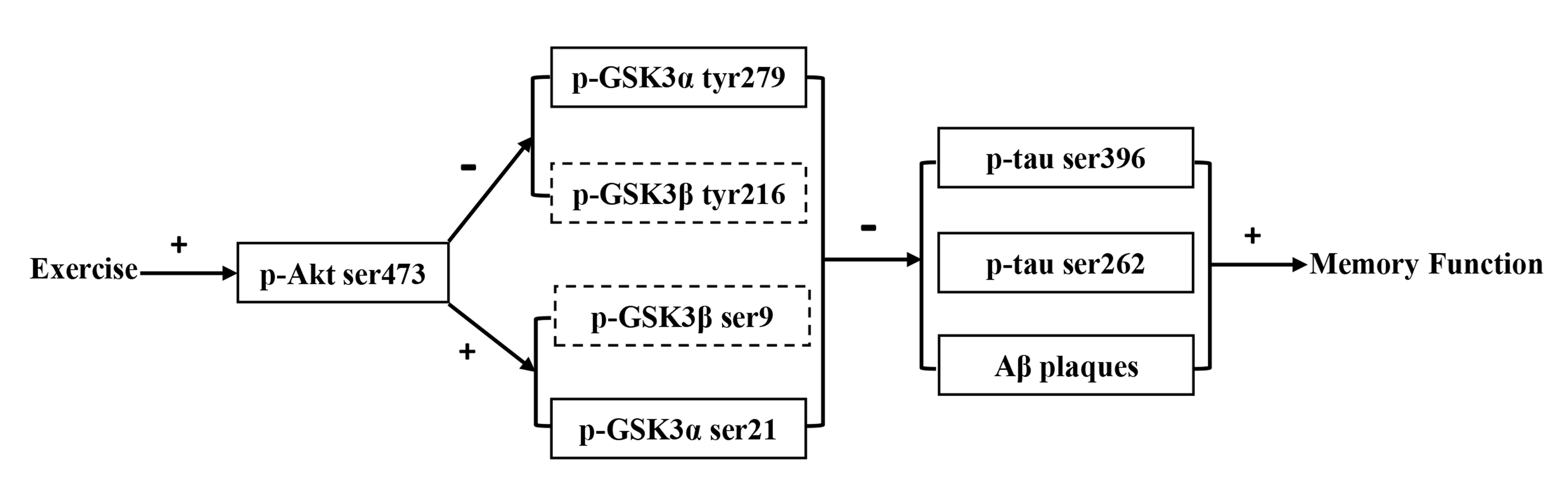 Fig. 7.
Fig. 7.
Schematic illustration of molecular reactions in the cortex and
hippocampus. Exercise induced the increased levels of p-Akt ser473 in both the
cortex and hippocampus, which subsequently led to increased inhibition of both
GSK3 and GSK3 in the two regions by increasing both
p-GSK3 ser21 and p-GSK3 ser9 but decreasing both
p-GSK3 tyr279 and p-GSK3 tyr216 in the hippocampus, as well as
by increasing p-GSK3 ser21 but decreasing p-GSK3 tyr279 in
the cortex. The inhibition of GSK3 and GSK3 in both regions
resulted in the reduction in both p-tau ser396 and p-tau ser262, as well as in
A plaques, and eventually improved memory function or delayed memory
deterioration in Tg-Ex mice. Note: solid lines indicate that the molecule was
present in both the cortex and hippocampus; dashed lines indicate that the
molecule was only present in the hippocampus. +, activation or improvement; –,
inhibition.
5. Conclusions
In conclusion, moderate-intensity aerobic training regulates
GSK3/ activity in the cortex and hippocampus of 9-month-old
APP/PS1 mice. The effects of exercise are more prominent in the hippocampus, but
similar in the cortex. As such, it is never too late to initiate an exercise
program to delay the progression of AD, which serves as a promising preventive
intervention.
6. Limitations
The present study used APP/PS1 Tg mice, which overexpress a chimeric
mouse/human APP (Mo/HuAPP695swe) and a mutant human presenilin 1 (PS1-dE9), both
of which exist in the central nervous system and are associated with early-onset
familial AD. Because this mouse model displays very similar biochemical aspects
to that of human AD, particularly regarding amyloid pathology, it is widely used
in studies of AD. However, it has inherent limitations with respect to the human
AD condition, as it represents only a small percentage of all AD patients. In
most AD patients, the disease is sporadic with a multifactorial etiology, making
it challenging to generalize findings obtained from this model to all AD
patients. Sporadic AD, the most common form, has not yet been faithfully modeled.
Nevertheless, the APP/PS1 model is a powerful tool in AD research.
Abbreviations
AD, Alzheimer’s disease; A, -amyloid; APP,
-amyloid precursor protein; GSK3, glycogen synthase kinase-3; NFTs,
neurofibrillary tangles.
Availability of Data and Materials
The datasets generated for this study are available
on request to the corresponding authors.
Author Contributions
PH and BG have contributed equally to this work. LZ and JY designed the research
study. PH, BG and LM performed the research. PH, BG and LM analyzed the data. PH and
BG wrote the manuscript. All authors contributed to editorial changes in the
manuscript. All authors read and approved the final manuscript. All authors have
participated sufficiently in the work and agreed to be accountable for all
aspects of the work.
Ethics Approval and Consent to Participate
The study was approved by the local ethics committee of Beijing Sport University
(2015015) and carried out according to the requirements of The Guiding Principles
for Care and Use of Animals.
Acknowledgment
The authors would like to gratefully thank the ethics committee of the Beijing
Sport University and Key Laboratory of Physical Fitness and Exercise of the
Beijing Sport University for supporting in this study.
Funding
This study was supported by National Natural Science Foundation of China
(31571229, 31271278) to LZ.
Conflict of Interest
The authors declare no conflict of interest.








 , Li Zhao 2,3,*
, Li Zhao 2,3,*