



Frontiers in Bioscience-Scholar (FBS) is published by IMR Press from Volume 13 Issue 1 (2021). Previous articles were published by another publisher on a subscription basis, and they are hosted by IMR Press on imrpress.com as a courtesy and upon agreement with Frontiers in Bioscience.
 , Satyan Lakshminrusimha 1
, Satyan Lakshminrusimha 11 Department of Pediatrics, Division of Neonatology, State University of New York at Buffalo, Buffalo, New York
2 Department of Pediatrics, University of California at Davis, Sacramento, California
Abstract
The etio-pathogenesis of necrotizing enterocolitis (NEC) is complex and multifactorial. Decades of research have not identified a definite etiology. Prematurity, enteral feeding, intestinal hypoxia/ischemia, inflammation and an abnormal microbiome are potential risk factors for developing this multisystem illness. Lack of specific diagnostic and prognostic markers adds to the challenges faced in managing NEC. Vascular mediators such as Nitric oxide (NO), catecholamines and endothelin (ET) regulate neonatal intestinal vascular resistance and may influence the pathophysiology of NEC. Neonatal morbidities, medications, transfusions, an altered microbiome and breast milk feeds may influence the vasculature in various ways. Better understanding of these mediators and their role in regulation of intestinal microcirculation and pathogenesis of NEC will assist in identifying strategies in prevention and management of this devastating illness.
Keywords
- Necrotizing Enterocolitis
- Nitric Oxide
- Endothelin
- Vasoconstriction
- Mesenteric Circulation
- Neonatal
- Review
The neonatal intestinal vasculature undergoes significant changes after birth as it transitions into an organ of nutrient absorption. It is regulated by a fine balance between vasodilator and vasoconstrictor forces. NO is the main vasodilator and endothelin-1(ET-1) the main vasoconstrictor maintaining neonatal intestinal vascular tone (1, 2). Any condition altering this balance has the potential to cause intestinal ischemia and injury. In this review, we will discuss the ontogeny of intestinal vascular regulation, the role of vascular mediators in NEC and factors that influence them. We will also discuss the beneficial effects of breast milk and potential future therapies for NEC that are currently being investigated.
In the fetus, the gastrointestinal tract is relatively dormant. Postnatally, it goes through a period of prolific growth and becomes a site of intense metabolic and anabolic activity. There is considerable lengthening of the small intestine from 125 cm at 20 weeks gestation to 275 cm at term and 380 cm at one year (3). Functionally, the intestines assume complete responsibility for assimilation of water and nutrients. The high oxygen demand required for these processes is met by maintaining high blood flow and oxygen delivery to the gut. The superior mesenteric artery (SMA) supplies blood to parts of the intestine derived from midgut (distal duodenum, jejunum, ileum, cecum and appendix, ascending colon, and proximal portion of transverse colon). Since the majority of intestinal blood flow relies on this single vessel, changes in SMA blood flow patterns may have significant physiological effects on the bowel.
Intestinal vascular resistance is high in fetal life and decreases postnatally (4, 5). This could be related to postnatal adaptation as well as the effect of enteral feeds (Table 1). 2-10 d old fed lambs have a higher intestinal blood flow and oxygen uptake as compared to fetal lambs (6). Ultrasound Doppler studies of the SMA in the perinatal period (fetal-30week, 2h, 24h and 72h) in healthy term infants show a similar pattern of increased intestinal perfusion, potentially to meet the increased metabolic demands of the bowel due to the onset of enteral feeding (7). Postnatal intestinal microcirculation in 3 d old swine is characterized by lower resting vascular resistance compared to that of older subjects (8). As the vascular resistance is inversely proportional to the vessel radius to the 4th power, small vasoconstrictive or vasodilatory changes can cause large changes in vascular resistance and blood flow (2, 9).
The regulation of gastrointestinal blood flow involves both intrinsic and extrinsic mechanisms acting at the tissue level (10). The myogenic component of intrinsic vasoregulation is the innate ability of the vascular smooth muscle to constrict in response to a stimulus. Regulation of tissue oxygenation based on metabolic demand constitutes the metabolic component which is mediated primarily by pre capillary sphincters and factors influencing arteriolar resistance. Reduction in arterial oxygen content decreases blood flow to the intestines and increases flow to brain and heart in fetal lambs through the “diving reflex” (11). The mechanism underlying such regulation at the level of the microcirculation is not well understood. Intestinal circulation in neonatal 3d old swine fails to autoregulate in response to reduced perfusion pressure (12). Reducing the flow rate in isolated perfused gut loops from 3d old swine for a sustained period causes an increase in vascular resistance (13). These findings suggest that reduced oxygen content and reduced perfusion pressure paradoxically further reduce blood flow to the intestines increasing the risk for ischemia.
Extrinsic vasoregulation has a neural and a humoral component. The neural component is mediated by the sympathetic nervous system and the humoral component by circulating vasoactive substances (10). Available experimental evidence suggests that under resting conditions, mucosal perfusion is mainly regulated by the vasodilators (NO and prostacyclin - PGI2), vasoconstrictors (endothelin) and by the sympathetic nervous system (14, 15) (Figure 1). A dynamic balance between vasoconstrictor and vasodilator forces exists in the intestinal vasculature. Vasoconstriction is mediated primarily by ET-1 and vasodilation by NO. In the newborn, this balance heavily favors vasodilation secondary to the copious production of endothelium-derived NO. Endothelial dysfunction occurring as a result of various factors alters this balance thus favoring ET-1-mediated vasoconstriction leading to ischemia and tissue injury (1).
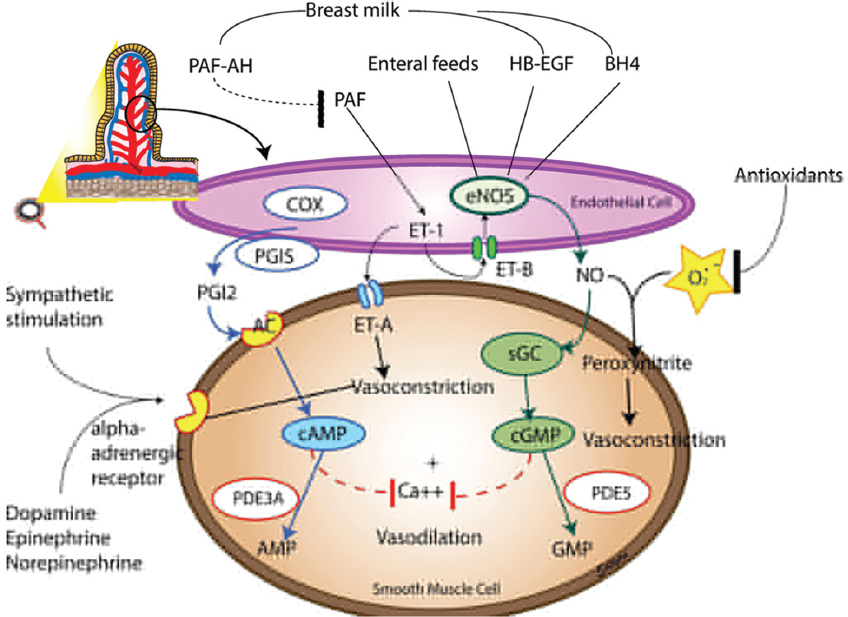 Figure 1
Figure 1Regulation of the intestinal vasculature. There is a dynamic balance between vasodilators and vasoconstrictors maintaining vascular tone and resistance. Vasodilation is mediated primarily by eNOS derived Nitric Oxide (NO) and prostacyclins. Several factors present in breast milk promote eNOS mediated vasodilation including platelet activating factor acetyl hydrolase (PAF-AH), heparin binding epidermal growth factor (HB-EGF) and tetrahydrobiopterin (BH4). The downstream pathway of vasodilation via NO includes activation of soluble guanyl cyclase (sGC) and cyclic guanosine monophosphate (cGMP). Cyclooxygenase (COX) and prostacyclin synthase (PGIS) are key enzymes involved in production of prostacyclins(PGI2) which activate the cyclic adenosine monophosphate (cAMP) pathway to produce vasodilation. Phosphodiesterase enzymes 3 and 5 (PDE3 and PDE5) degrade cAMP and cGMP respectively. Endothelin 1(ET-1), acting through ET-A receptors and catecholamines acting on the adrenergic receptors act as the primary vasoconstrictors. In addition, reactive oxygen species produced as a result of various metabolic pathways may combine with NO to produce peroxynitrite and cause vasoconstriction. ET-B receptors promote vasodilation through the production of NO, however the vasoconstrictive action through ET-A exceeds the vasodilatory effect through ET-B causing ET-1 to have a net vasoconstrictive effect.
NO is produced by the enzyme nitric oxide synthase which has three isoforms- Endothelial nitric oxide synthase (eNOS), inducible nitric oxide synthase (iNOS) and neuronal nitric oxide synthase (nNOS). eNOS and iNOS play a more significant role in maintaining NO in mesenteric vasculature. iNOS is upregulated in certain conditions including inflammation and produces high levels of NO which may contribute to the deleterious effects of NO products such as peroxynitrite. These effects will be discussed later in this review. eNOS is constitutively expressed in the intestinal vasculature and is responsible for the low background levels of NO. This eNOS mediated NO production is thought to contribute to maintenance of vascular tone through the cyclic guanosine monophosphate (cGMP) pathway.
eNOS-derived NO is the most potent vasodilator stimulus present in the newborn intestine(16). NO produced in the endothelium diffuses to adjacent vascular smooth muscle cells, where it binds to the heme moiety of soluble guanylate cyclase (sGC), activating sGC to produce cGMP(1)(Figure 1). Most studies evaluating the role of endothelium derived NO have been performed in sheep and swine. Endogenous NO has been shown to be a significant contributor to the modulation of vascular tone in the GI circulation of mid as well as late-gestation fetal lambs (17, 18) http://www.nature.com/pr/journal/v44/n3/full/pr19982237a.html - bib6. Postnatally, this role of nitric oxide is age dependent, being more prominent in 3 d old as compared to 35d old piglets (9). Though abundant eNOS mRNA is present in fetal and postnatal mesenteric arteries (19), eNOS protein peaks at term gestation and remains high in postnatal ovine mesenteric arteries, exhibiting a distinct ontogeny (20).
Several factors may upregulate this mechanism. Flow regulated vasodilation in the intestinal circulation is mediated by eNOS derived NO and is more prominent in the immediate postnatal period (21). Enteral feeds can induce a post prandial hyperemia in splanchnic circulation, the exact etiology of which is not well understood. Doppler ultrasound studies of the mesenteric vessels in preterm infants have shown a post prandial increase in mesenteric flow after the first feed (22). Reber et al noted increased eNOS protein in mesenteric arteries of 1-d-old fed piglets compared with fetal and 1-d-old unfed piglets, suggesting that there may be an upregulation of this enzyme after enteral feeds (19). Other factors that may affect eNOS expression and activity are proinflammatory cytokines, such as tumor necrosis factor (TNF) (23), interleukin-1(IL-1)(24) and toll like receptor 4 (TLR 4) (25). Heparin-binding epidermal growth factor-like growth factor (HB-EGF) is a member of the epidermal growth factor family that has been studied in NEC. HB-EGF also stimulates eNOS protein production and NO release from cells (26).
ET-1 is thought to be the primary constrictor stimulus in the intestinal circulation (27) and implicated in intestinal vascular dysfunction and tissue injury. While it exerts its action through receptors A and B (ETRA and ETRB), it has been shown that the force of ETRA-induced vasoconstriction exceeds that of ETRB-generated vasodilation (1). Local, constitutive production of ET-1 helps in establishing basal vascular tone in terminal mesenteric arteries from 1 to 5 day old swine via activation of both ETRA and ETRB receptors (27, 28). ET-1 production can be stimulated by decreased blood flow, hypoxia and various inflammatory cytokines (29, 30).
ET-1 has an age-dependent effect within postnatal intestine, being more prominent in 3 d old as compared to 35 d old swine.(27) Researchers have demonstrated a profound reduction of intestinal perfusion and subsequent tissue damage following intra-arterial infusion of ET-1 (31, 32). Moreover, ET-1 participates in the generation of sustained intestinal ischemia and subsequent tissue damage following a single episode of ischemia-reperfusion (33).
Other vasoconstrictor mediators in newborn intestine include angiotensin (ANG) and catecholamines like norepinephrine (NE). Catecholamines exert their vasoconstrictive effects on circulation via alpha adrenergic receptors. Alpha adrenergic receptor 1A (ADRA1A) appears to be the predominant receptor in mesenteric circulation in rat, dog and humans (34-36). Sustained reduction of blood flow in newborn swine intestine decreases constitutive NO production and subsequently enhances the contractile efficacy of vasoconstrictors ANG II, NE and ET-1 (13). In our studies on ontogeny of ovine mesenteric vasculature, we have shown that mesenteric arterial constriction to NE peaks at 134 d corresponding to late preterm gestation (Figure 2). In addition, ADRA1A mRNA and protein increased significantly at 134d and decreased postnatally, suggesting that mesenteric vasoconstrictive forces predominate in late-preterm gestation (20).
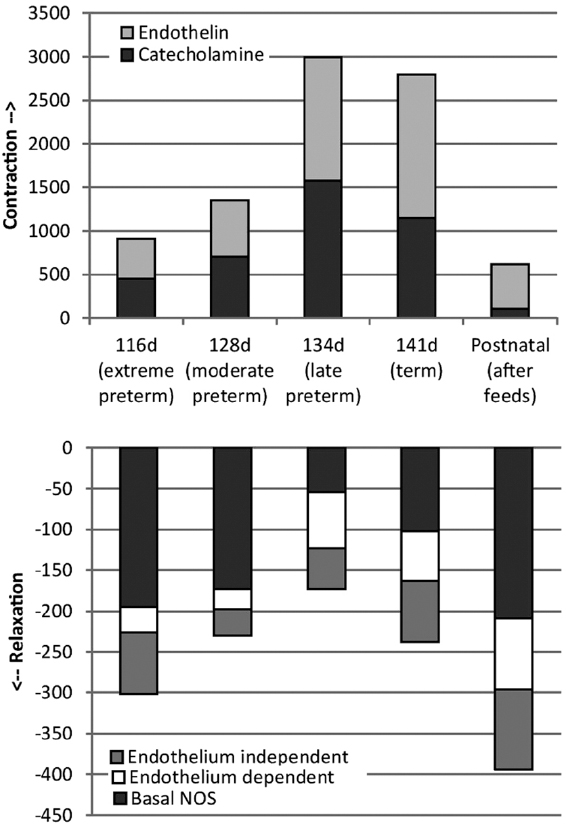 Figure 2
Figure 2Ontogeny of mesenteric arterial contractility. Distinct ontogenic patterns are noted in mesenteric arterial constriction and relaxation patterns in ovine fetal and postnatal mesenteric arteries. The vasoconstrictive ability of mesenteric arteries in response to ET-1 and Norepinephrine (NE) peaks at 134d (late preterm) gestation and decreases postnatally after enteral feeds. In contrast, the mesenteric arteries of fetal lambs achieve maximal relaxation after birth and enteral feeds. 134d gestation equated to late preterm gestation in humans is characterized by maximal constriction and least relaxation responses. (Adapted from Nair et al, Fetal and postnatal ovine mesenteric vascular reactivity. Pediatr Res. 2016;79(4):575-582).
Abnormalities of intestinal circulation have been observed in NEC. High SMA vascular resistance is noted in preterm babies who later developed NEC (37). Coagulation necrosis, which is characteristic of ischemic injury, was noted in tissue samples of infants with NEC (38). Ischemia and reperfusion injury (I/R) has long been postulated as one of the etiological factors in NEC with preterm neonates being more susceptible to multiple episodes of low flow states, hypotension and hypoxia. Animal models of NEC have utilized hypoxia/reoxygenation to experimentally induce NEC (39). However, risk factors like intrapartum asphyxia, patent ductus arteriosus and umbilical artery catheters are associated with equal frequency in preterm infants with and without NEC (40). This has led to the current presumption that though hypoxia and ischemia are unlikely to be the primary initiating event in preterm infant, the regulation of intestinal circulation, especially the microcirculation, does play a role in NEC pathogenesis.
However, in term infants, a vascular etiology is often identified. Congenital left sided heart disease, gastrointestinal abnormalities, hypoxic ischemic injury as well as polycythemia are various risk factors in term infants that are associated with the development of this process (41). Imbalance in the vasoconstrictor vasodilator balance may contribute to this disease process. Nowicki et al show that while eNOS expression is maintained, eNOS activity and NO production are decreased in mesenteric arterioles isolated from infants with NEC(16). Animal models of NEC have yielded useful information in the role of NO pathway in this condition. NOS blockade decreases intestinal microcirculatory blood flow in a rat model of NEC (42). In human preterm infants with NEC, increased tissue concentration of ET-1 and increased arteriolar vasoconstriction is noted in intestinal samples with evidence of NEC as compared to those from relatively healthy intestine in the same specimen (43). Blockade of ETRA receptors reversed these effects.
Several studies have noted an inverse relation with gestational age with the most premature infants having the highest risk of developing NEC (44, 45). NEC is rare in term infants, but when it occurs, it presents in the first week of life. NEC is common in extremely premature infants but presents after a few weeks of postnatal life. The median postmenstrual age at NEC presentation is usually around 31-34 weeks (44-46). Some studies have suggested that a unique factor predisposes preterm infants to develop NEC at this postmenstrual age (1). We have previously shown that ovine mesenteric vasoconstriction mediated by endothelin and catecholamines predominates in late-preterm gestation followed by a conversion to vasodilatory influences, mediated by NO, near the time of full-term birth (20)(Figure 2). Distinct ontogenic patterns in vascular reactivity have been noted in chick embryo where maturation of vasodilator mechanisms preceded that of vasoconstrictor mechanisms (47).
While NO derived from vascular endothelium (mainly through eNOS) promotes vasodilation and is protective against NEC, NO derived from other sources – mainly iNOS may play a role in the pathogenesis of NEC. These effects are mediated through intestinal oxygen consumption and/or disruption of the epithelial barrier permitting invasion by an abnormal microbiome.
NO has a direct effect on cellular respiration-it reversibly inactivates cytochrome c within the mitochondrial electron transport chain (48). Nowicki et al have shown that iNOS-derived NO suppressed intestinal oxygen consumption in intestine resected for NEC (49). Thus, suppression of intestinal oxygen consumption is yet one more means whereby iNOS-derived NO contributes to the pathogenesis of NEC.
The pathogenesis of NEC involves disruption of the intestinal epithelial barrier, due to a combination of local and systemic insults (50). NO is thought to play an important role in the intestinal barrier failure seen in NEC. In high concentrations it reacts with superoxide to form its reactive nitrogen derivative peroxynitrite. NO and peroxynitrite may affect gut barrier permeability by inducing enterocyte apoptosis and necrosis(50), or by altering tight junctions or gap junctions that normally play a key role in maintaining epithelial monolayer integrity (51). NO and metabolites may also inhibit the epithelial restitution processes, both enterocyte proliferation and migration (52). While iNOS plays a major role in NO production here, factors that govern iNOS upregulation in the intestine are not well understood. Increased iNOS expression has been noted in intestinal epithelial cells in surgically resected tissue samples of NEC (53). In a rat model of NEC, induced by formula feeding, upregulation of iNOS mRNA, enterocyte apoptosis, and decreased IL-12 production in the intestinal epithelium was noted (54). High levels of NO can lead to increased bacterial translocation by their detrimental effects on the gut barrier (51).
The role of the intestinal microbiome in the pathogenesis of NEC is still being investigated. NEC does not occur in germ free animals (55, 56), however, a definite pathogenic relationship with specific organisms is not seen. It is possible that dysbiosis or altered microbiome plays a role, interacting with an already immature intestine to contribute to NEC. The intestinal microbiome and bacterial colonization begins to get established in the perinatal period. Organisms that colonize the intestines and their components can induce production of NO mediated by iNOS. Infection with Cronobacterium or E. sakazakii induced NEC in a newborn rat model by inducing enterocyte apoptosis (57). Furthermore, this organism in rat intestinal epithelial cells (IEC-6) induced significant production of nitric oxide (NO). This elevated production of NO due to increased expression of iNOS was responsible for apoptosis of IEC-6 cells (58). Better understanding of the role of NO and iNOS induction are necessary for developing iNOS-targeted NEC therapies (52).
The final common pathway of NEC involves production of inflammatory mediators induced by ischemia, infectious agents, or mucosal irritants which recruit inflammatory cells and further contribute to tissue injury (59). Some mediators that have been recognized for their role in NEC pathogenesis include TNF, interleukins (IL-1, IL-6, IL-8, IL-10, IL-12, and IL-18) and platelet activating factor (PAF) (60). Preterm infants with NEC show high levels of these cytokines. There is some correlation between levels of cytokines and extent or severity of disease (61, 62) (63).
Some cytokines play a protective role. IL-10 is present in preterm breast milk and a paucity of IL-10 in human milk was found in mothers whose infants developed NEC(64). The mechanism of action is not known. However, intraperitoneal administration of IL-10 has been shown to decrease the severity of intestinal injury and the level of NO synthesized by inhibiting iNOS expression in an IL10 knockout rodent NEC model (65).
Inflammatory mediators also seem to influence vascular tone by interacting with the vasoconstrictors and vasodilators mentioned earlier in this review. PAF, synthesized by many inflammatory cells and bacteria, can cause intestinal necrosis in animals (66). This agent can increase transcription of the vasoconstrictor ET-1 receptor which in turn can further activate PAF (67). Human infants with NEC have increased levels of PAF and decreased expression of PAF acetylhydrolase (PAF-AH), which is the enzyme responsible for degrading PAF (68). Human milk contains PAF-AH but this compound is absent in infant formula. Enteral administration of the recombinant PAF-AH enzyme markedly reduced the incidence of NEC in a rat model of the disease (69).
Oxidative stress is defined as an imbalance between production of oxidative reactive species and antioxidant defenses. Premature infants, in general, have poor antioxidant defenses since the physiologic increase in antioxidant capacity occurs at the end of gestation (70). Oxidative stress (71) and reactive oxidative species like superoxide have been implicated in NEC pathogenesis. This may involve components of the NO, NADPH oxidase (72) and xanthine oxidase pathway. Certain conditions including tetrahydrobiopterin deficiency can cause uncoupling of eNOS, resulting in a switch from producing NO to superoxide (73). In neonatal rat pups, intestinal ischemia is associated with this NOS dependent uncoupling mechanism causing over-production of reactive oxygen species and impaired vasodilation (74). In addition, stress induces iNOS causing high concentrations of NO in the intestinal microcirculation (74, 75). This NO may react with available superoxide to produce peroxynitrite, another oxidant agent that causes further cellular and tissue damage(76). Magnesium and copper deficiency impair antioxidant defense through decreased synthesis of glutathione and reduced activity of Cu/Zn superoxide dismutase (77). Recently there is some evidence to suggest that maternal magnesium sulfate reduces cytokine IL-6 and TNF-α production in neonates, suggesting potential as therapy in various neonatal inflammatory conditions (78). The role of antioxidants like all-trans-retinoic acid (ATRA) (79) and N-acetylcysteine (NAC)(80) in treating NEC is being investigated in animal models. Enteral glutamine and/or arginine (precursors of NO) supplementation have shown favorable effects on oxidative stress parameters in neonatal rat intestine (81).
Over the last few years, several studies noted an association between packed red blood cell (PRBC) transfusions and the development of NEC (82-84). “TRAGI” or transfusion associated gut injury, the term proposed by Blau et al (83), was thought to be due to a failure of the postprandial increase in mesenteric blood flow following a transfusion.
Blood products affect arterial reactivity and intestinal motility (85). Beneficial effects of transfusion include hypoxic vasodilation, proposed to be due to the action of NO through S-Nitrosylation of hemoglobin (SNO-Hb). Within red blood cells (RBCs), Hb is S-nitrosylated during the respiratory cycle and thereby conveys NO bioactivity that may be dispensed to regulate local blood flow in the physiologic response known as hypoxic vasodilation. Hb thus both delivers oxygen directly and delivers vasoactivity to potentially optimize tissue perfusion in concert with local metabolic demand. It has been shown recently that storage of blood depletes S-nitrosylated Hb(86), accompanied by reduced ability of RBCs to induce vasodilation. This “storage lesion” appears to account in significant part for the impaired ability of banked RBCs to deliver oxygen. The mechanisms involved include reduced RBC deformability, increased red blood cell adhesion and aggregation, prothrombotic tendency and nitric oxide deficiency(87). Renitrosylation can correct this impairment and thus may offer a means to ameliorate the disruptions in tissue perfusion produced by transfusion(86). PRBC transfusion in enterally fed preterm lambs promotes mesenteric vasoconstriction and impairs vasorelaxation by reducing mesenteric arterial eNOS (88). In addition, recently, investigators have found that PRBC have the potential to release high quantities of the endogenous NOS inhibitors ADMA and LNMMA (89). RBC transfusion can also significantly increase post-transfusion plasma free Hb levels and plasma NO consumption in the recipient (90).
We have evaluated the changes in oxygen extraction and SMA blood flow in preterm lambs (134 d gestation; term ~ 147 d) with anemia induced by partial exchange transfusion (91). Preterm lambs with normal hematocrit had 21.8.7±6.6.5 ml/kg/min of SMA flow and an oxygen extraction ratio (OER) percentage of 9.3.7±1.5. Enteral feed with ewe’s colostrum increased SMA flow and increased oxygen delivery to the intestines (Figure 3). The OER stayed the same. Lambs with anemia had a higher SMA flow at baseline to compensate for lower oxygen content in the blood. Following colostrum feed, SMA blood flow did not further increase but the oxygen demand was met by increased oxygen extraction (Figure 3). Thus, the increased oxygen demand in response to enteral feeding is met by increasing blood flow and/or oxygen extraction. Transfusion of PRBC (7 days old) was followed by an enteral feed. A prior transfusion prevented postprandial increase in intestinal blood flow and oxygen extraction. In the presence of severe anemia and low oxygen delivery to the intestine, we speculate that transfusion followed by feeds increases oxygen demand above oxygen supply leading to ischemia.
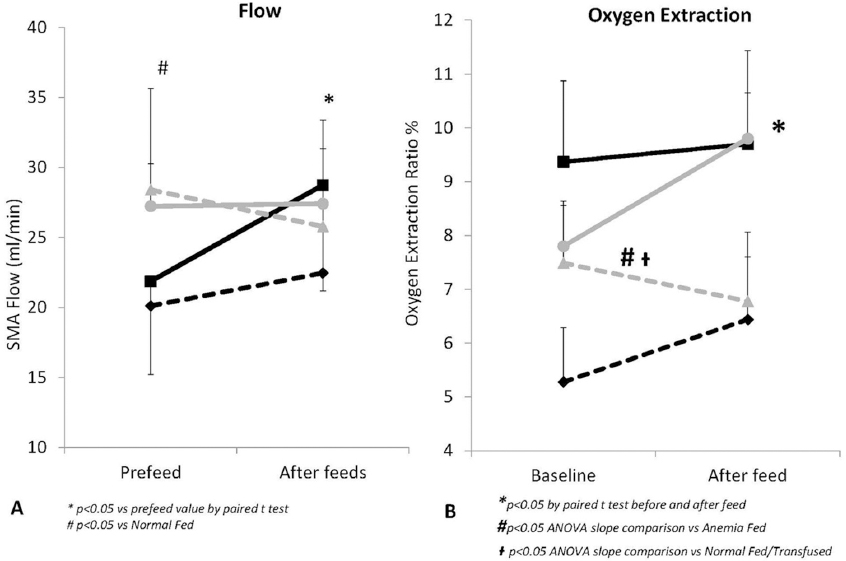 Figure 3
Figure 3Mesenteric arterial flow (A) and oxygen extraction (B) are affected by enteral feeds and transfusions in preterm lambs. After enteral feeds, SMA flow rises significantly in normal preterm lambs (black solid line) without any significant change in oxygen extraction ratio. Transfusion with pRBC (black dashed line) blunts this postprandial rise in flow. Anemic preterm lambs (grey solid line) have a higher baseline SMA flow which does not show any significant change after enteral feeds. Instead, oxygen extraction rises after feed to meet tissue metabolic needs. Transfusion with pRBC in these lambs (grey dashed line) does not change the SMA flow but significantly decreases oxygen extraction as well. (Adapted from Nair et al, Transfusion Decreases Intestinal Oxygen Extraction in Anemic Preterm Lambs After Feeds Pediatric Academic Societies Abstracts, 1705.8. 2014) Figure key: Black solid line- normal preterm lambs, black dashed line- normal preterm lambs transfused with pRBC, grey solid line- anemic preterm lambs, grey dashed line- anemic preterm lambs transfused with pRBC.
More recently, the consensus on the etiopathogenesis of transfusion associated NEC is that the state of chronic anemia contributes significantly to this condition (92). In a large multicenter study in very low birth weight (VLBW) infants, severe anemia, but not RBC transfusion, was associated with an increased risk of NEC (93). Krimmel et al (94) looked at doppler SMA flow velocities and noted that if bolus feedings were preceded by blood transfusions, only anemic neonates weighing <1250 grams failed to develop a postprandial rise in mesenteric artery blood flow velocity. Lower hemoglobin levels may cause gut injury though interactions between hypoxemia, enteric blood flow and feeding. A trial evaluating optimal threshold for transfusion of preterm infants (TOP) is currently recruiting patients and will assess the incidence of NEC in higher and lower hemtocrit threshold groups of preterm infants (Clinical trials no NCT01702805).
VLBW infants are prone to several morbidities as a result of their gestational age and immature adaptation abilities. Physiological instability, mechanical ventilation, increased hemodynamic support or extensive neonatal resuscitation and hypotension are reported as additional risk factors for NEC (95). In addition, the use of vasopressors in the first week of life has been shown to be associated with mortality in infants who subsequently developed NEC(96). This likely signifies the extent of their hemodynamic instability and hypotension which contributed to the disease process. One of the most common medications used to manage hypotension is dopamine. However this medication itself may cause significant effects on intestinal vasculature. Dopamine caused smooth muscle contraction in mesenteric arterioles and precapillary sphincters of anesthetized dogs, producing intestinal ischemia and hypoxia (97). While doppler studies in preterm infants have shown that dopamine increased mesenteric blood flow velocity and decreased mesenteric vascular resistance (98), dopamine-induced increases in SMA blood flow have been associated with decreases in oxygen extraction and mucosal blood flow (99, 100).
A patent ductus arteriosus (PDA) can be a risk factor for NEC (101),(102). Due to the systemic steal phenomenon, there is potentially decreased mesenteric perfusion in a hemodynamically significant PDA, predisposing the infant to intestinal ischemia. Preterm infants with PDA have been reported to have low mesenteric blood flow velocity (103). Indomethacin, ibuprofen and more recently acetaminophen are medications commonly used in the NICU to treat hemodynamically significant PDAs. Indomethacin, a cyclooxygenase (COX) inhibitor, independently reduces intestinal blood flow (104, 105) and has been associated with increased incidence of NEC in VLBW infants (106, 107). However other large studies like the Trial of Indomethacin Prophylaxis in Preterm Infants (TIPP) did not find a significant increase in the risk of NEC with indomethacin as prophylaxis for intraventricular hemorrhage or PDA (108) and it still continues to be used to treat this condition. Ibuprofen does not decrease mesenteric blood flow (105) and may a better pharmacologic choice to treat a PDA without increasing the risk of NEC (109).
Breast milk is known to be protective in NEC (110, 111). Besides immunological factors like secretory IgA and anti-inflammatory cytokines that are present in breast milk, there are several other compounds such as erythropoietin and oligosaccharides that have been studied in relation to NEC. Breastfeeding also influences the composition of infant’s gut microbiota(112). EGF found in breast milk, has been noted to protect intestinal epithelium against injury (113). Administration of exogenous HB-EGF in rodents protects the intestines from experimental NEC (114). Tetrahydropterin (BH4) present in breast milk is a required cofactor for eNOS coupling and NO generation and has been shown to enhance mesenteric blood flow (115). Exposure to BH4 reduces newborn rat mesenteric arterial vascular tone (116). Higher biopterin concentrations have been observed in human breast milk, when compared with infant formula (116). This may potentially be one of the factors that explain the protective effects of breast milk over formula use in NEC patients.
NO is synthetized from the conditionally essential amino acid L-arginine by NO synthases (117). Researchers found that continuous intravenous infusion with L-arginine markedly reduced intestinal injury in a neonatal pig model of NEC (118). Premature infants with NEC were noted to have decreased plasma arginine concentrations at the time of diagnosis (119). The role of L-arginine supplementation has been explored in small studies and shows some promise in reducing the incidence of NEC (120, 121). However, larger studies are required to assess its efficacy in preventing NEC in VLBW preterm neonates before widespread use can be recommended.
Inhaled NO exerts beneficial effects on ischemia/reperfusion injury in extra pulmonary organs in animal (122) as well as human studies (123, 124). Administration of inhaled NO has been shown to reduce the size of infarct following stroke in rodent models (125). A similar approach to NEC has been proposed and is being currently investigated in preterm infants (Clinical trials # NCT02851472).
Similar to the lung, the fetal intestine is a relatively dormant organ whose nutritive function is performed by the placenta. Just as breathing air results in pulmonary vasodilation, enteral feeding reduces intestinal vascular resistance. The intestinal oxygen demand increases with enteral feeding and inability to meet this demand due to stressors such as severe anemia, prematurity and transfusions causing a predominant vasoconstrictor stimulus may lead to disruption of the epithelial barrier. Finally, the complex interaction between intestinal microbiome, epithelial barrier and vasodilators such as NO may play a role in the pathogenesis of NEC. Understanding these interactions may enable us to come up with newer, innovative, preventive and therapeutic strategies against NEC.