
 , Qiang Wen 2, Haigang Ding 1, Jiangjing Shan 1, Qi Zhang 3,*
, Qiang Wen 2, Haigang Ding 1, Jiangjing Shan 1, Qi Zhang 3,*
1 Department of Gynecology, Shaoxing People’s Hospital (The First Affiliated Hospital, Shaoxing University), 312000 Shaoxing, Zhejiang, China
2 Department of Gynecological Oncology, Zhejiang Cancer Hospital, 310022 Hangzhou, Zhejiang, China
3 Department of Ultrasound, Shaoxing People’s Hospital (The First Affiliated Hospital, Shaoxing University), 312000 Shaoxing, Zhejiang, China
Abstract
This review provides a comprehensive analysis of copper metabolism and cuproptosis in ovarian cancer (OC), evaluating therapeutic strategies targeting copper to improve clinical outcomes.
OC exhibits the highest mortality rate among gynecological malignancies, characterized by its insidious onset and poor prognosis, underscoring the urgent need for novel treatment strategies. Copper metabolism and cuproptosis hold promising potential in regulating tumor progression and overcoming drug resistance, thereby opening new avenues for OC treatment. This article aims to elucidate the biological basis and potential therapeutic applications of copper metabolism and cuproptosis in treating OC.
Cuproptosis, a copper-dependent mechanism of cell death mechanism, disrupts mitochondrial function by inducing the aggregation of lipoylated proteins and the loss of iron-sulfur (Fe-S) clusters. Copper metabolism imbalance promotes OC progression by modulating cancer cell function, contributing to chemotherapy resistance, and influencing responses to anti-angiogenesis, and immunotherapy. Copper ionophores, chelators, copper-based nanoparticles (NPs), and certain natural molecules represent potential therapeutic strategies for the treatment of OC.
Targeting copper metabolism and cuproptosis offers innovative therapeutic strategies for OC, particularly in cases of treatment resistance. However, clinical validation of long-term safety, optimal dosing, and biomarker-guided therapies remains critical. Future research should prioritize translational studies to bridge mechanistic insights with patient-centered applications.
Keywords
- copper metabolism
- cuproptosis
- ovarian cancer
- review
Due to the absence of specific detection methods, approximately 70% of ovarian cancer (OC) cases are diagnosed at an advanced stage, leading to a poor prognosis [1]. According to global cancer statistics, OC accounted for 324,398 new cases and 206,839 deaths in 2022 [2]. Clinical treatments for OC include surgery, combination chemotherapy, and targeted maintenance therapy. In recent years, the introduction of poly (ADP-ribose) polymerase inhibitors (PARPis) has led to significant advancements in OC treatment [3]. Nevertheless, the overall cure rate remains low. Many patients experience relapse within 18 months and ultimately succumb to chemotherapy resistance, with a 5-year survival rate of only 49% for those diagnosed at advanced stages [4]. Chemotherapy and PARPis have been associated with several adverse drug effects, including bone marrow suppression and gastrointestinal reactions. Therefore, it is crucial to explore novel and safe treatment strategies for OC to enhance the prognosis of patients with this disease.
As an essential trace element, copper plays a catalytic or auxiliary role in various biological processes. Serum copper levels are significantly elevated in patients with OC, suggesting its potential as a biomarker for this disease [5, 6]. The bioavailability of copper and the regulation of cellular homeostasis influence cell proliferation, migration, and angiogenesis, processes that are critical in the progression and chemoresistance of OC [7, 8]. A novel form of cell death, termed “cuproptosis”, was introduced in March 2022 as a groundbreaking concept [9]. This copper-dependent and modifiable form of cell death may help reduce drug resistance in cancer therapy and holds significant promise for cancer prevention and treatment [10]. Cuproptosis-related proteins have been identified as key players in OC [11]. Despite advancements in understanding the roles of copper metabolism and cuproptosis in OC, the specific mechanisms remain insufficiently elucidated. Leveraging the mechanisms of copper metabolism and cuproptosis to develop copper ionophores, copper chelators, or copper-based nanoparticles (NPs) for the selective induction of cell death in OC may offer a novel therapeutic strategy for the disease.
In this study, we present a comprehensive review and analysis of the mechanisms underlying copper metabolism and cuproptosis in OC. The primary goal is to elucidate their regulatory roles in tumor progression and explore their potential applications in OC treatment. Through comprehensive research, we aim to strengthen the theoretical foundation and practical implications of copper metabolism and cuproptosis in OC therapy, providing valuable insights for the development of effective treatment strategies.
Copper is an essential trace nutrient for normal organ function and metabolic
processes in animals and humans. The body primarily acquires copper from food;
however, dietary copper in the form of copper (II) (Cu2+) cannot be directly
utilized by cells. Dietary copper absorption primarily occurs in the small
intestine and relies on copper transporter 1 (CTR1), located on the apical
membrane of intestinal epithelial cells [12]. CTR1, a high-affinity copper
importer from the Solute Carrier Family 31 (SLC31), is crucial for copper
homeostasis. Six-transmembrane epithelial antigen of the prostate (STEAP) and
duodenal cytochrome B (DCYTB) play a crucial role in the reduction of Cu2+
to Cu+, facilitating the subsequent transport of copper into intestinal
epithelial cells via CTR1 [13, 14]. Once inside the intestinal epithelial cells,
Cu+ is released into the portal circulation by the copper-transporting
ATPase
The digestive tract is the primary route for copper excretion, with approximately 80% of endogenous copper excreted into the gastrointestinal tract via bile. A small amount of this copper can be reabsorbed with saliva, gastric acid, and intestinal fluid, while the remainder is excreted in the feces [22]. Copper that enters the gastrointestinal tract through food and is not absorbed, referred to as exogenous copper, is excreted in the feces. While the primary route for endogenous copper excretion is via bile, other excretion pathways include urine, sweat, and menstruation [15].
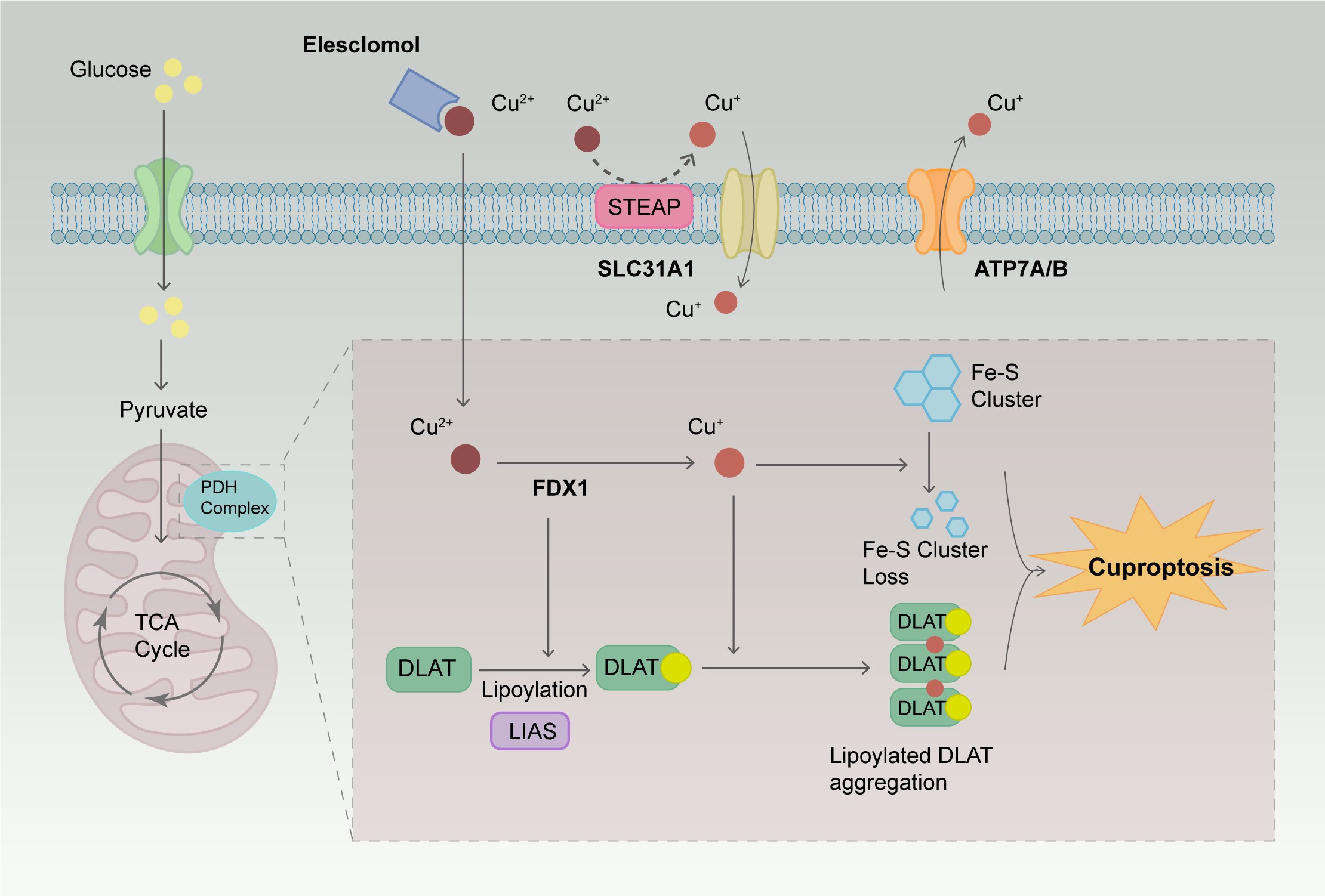
In 2019, researchers discovered two small molecules, disulfiram (DSF) and elesclomol (ES), which can transport copper ions across cell membranes [23]. They referred to these molecules as copper ionophores, important tools for studying copper-induced cell death [23]. Research has shown that cells with a high dependency on mitochondrial respiration are nearly 1000 times more sensitive to copper ionophores than cells relying on glycolysis [23]. There is a significant difference in the sensitivity of cells treated with mitochondrial function inhibitors compared to those treated with ferroptosis inhibitors targeting glutathione peroxidase-4 (GPX4). Treatment of cells with the copper ionophore ES disrupts the metabolism of tricarboxylic acid (TCA) cycle-related products, further elucidating the close connection between copper ionophore-induced cell death and the TCA cycle. This copper-induced cell death process was first named cuproptosis in 2022. The primary mechanism of cuproptosis depends on the accumulation of copper ions within the cells. When an excess amount of Cu2+ enters the cells, it is transported to the mitochondria and reduced to Cu+. This Cu+ interferes with the TCA cycle and the electron transport chain (ETC), leading to oligomerization of lipoylated proteins and loss of iron-sulfur (Fe-S) cluster proteins, ultimately resulting in cell death. To further investigate the metabolic pathways of cuproptosis, Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated protein 9 (Cas9) technologies were used for loss-of-function screening, leading to the identification of 10 key genes regulating cuproptosis activity [8]. The mechanisms of these genes are presented in Table 1 [9, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57]. Lipoylation plays a vital role in maintaining cellular metabolic pathways and normal mitochondrial function [9]. Ferredoxin 1 (FDX1)/lipoyl synthase (LIAS) are key upstream regulators of protein lipoylation and play a crucial role in regulating cuproptosis in cells. FDX1 possesses strong reducing capabilities, which enable it to reduce Cu2+ to Cu+ and lipoylate specific enzymes in the mitochondria, such as dihydrolipoamide S-acetyltransferase (DLAT), glycine cleavage system protein H, dihydrolipoamide S-succinyltransferase, and dihydrolipoamide branched chain transacylase E2 [9]. ES can bind Cu2+ outside the cell and transport it into the cell. Once inside the cell, Cu2+ is reduced by FDX1 to Cu+, which can bind to lipoylated proteins, causing protein aggregation and disrupting the structure, function, and activity of enzymes, thereby interfering with the normal function of the TCA cycle. Cu+ also affects the function of Fe-S cluster proteins in the mitochondria, including electron transfer, catalytic reactions, and DNA repair [58]. The mechanism of cuproptosis is illustrated in Fig. 1.
| Genes | Regulation of cuproptosis | Mechanism | References |
| FDX1 | (+) | It facilitates electron transfer from NADPH to mitochondrial cytochrome P450. The copper ionophore binding to FDX1 can upregulate the accumulation of intracellular copper ions and promote the death of OC cells. It is associated with RNA methylation modifications. It is involved in cisplatin resistance in OC. | [24, 25, 26] |
| LIPT1 | (+) | It is an essential enzyme for the activation of mitochondrial 2-ketoacid dehydrogenase, participating in fatty acylation. It can regulate lipoic acid metabolism. Its overexpression is associated with a favorable prognosis in OC patients. | [27, 28, 29] |
| LIAS | (+) | It participates in the synthesis of antioxidant lipoic acid in mitochondria and is closely related to mitochondrial energy metabolism and oxidative stress. Its high expression is related to the good prognosis in patients with OC. | [30, 31, 32, 33] |
| DLD | (+) | It is a key protein of the TCA cycle and constitutes the E3 component of the |
[34, 35] |
| DLAT | (+) | It catalyzes the decarboxylation of pyruvate to acetyl-CoA in the TCA cycle. It affects the energy supply of OC cells. | [9, 36, 37, 38, 39] |
| PDHA1 | (+) | It catalyzes pyruvate decarboxylation and serves as a gate-keeper enzyme link between glycolysis and the mitochondrial TCA cycle. Its decreased expression predicts an unfavorable prognosis in OC. Its low expression is related to cisplatin resistance in OC. | [40, 41, 42] |
| PDHB | (+) | It catalyzes the conversion of glucose derived pyruvate to acetyl-CoA, thereby regulating oxidative phosphorylation. It serves as a glycolysis regulatory gene that can play a role in the development of cancers, including OC. It is upregulated in OC. | [43, 44, 45, 46] |
| MTF1 | (–) | As a protein-coding gene, it can activate the transcription of MTs gene by binding to MREs. It is upregulated in OC, and its high expression is associated with poor patient survival and disease relapse. | [47, 48, 49] |
| GLS | (–) | It catalyzes the conversion of glutamine to glutamate. Knockdown of glutaminase in platinum-resistant cells results in re-sensitization to platinum treatment. | [50, 51, 52, 53] |
| CDKN2A | (–) | Also known as p16, it is a tumor suppressor protein involved in regulating the cell cycle at the level of transition from G1 to S phase. Its mutation and promoter methylation are associated with OC. Its mutation is related to the recurrence of OC. | [54, 55, 56, 57] |
FDX1, ferredoxin 1; LIPT1, lipoyltransferase 1; LIAS, lipoyl synthase; DLD,
dihydrolipoamide dehydrogenase; DLAT, dihydrolipoamide S-acetyltransferase;
PDHA1, pyruvate dehydrogenase E1 subunit
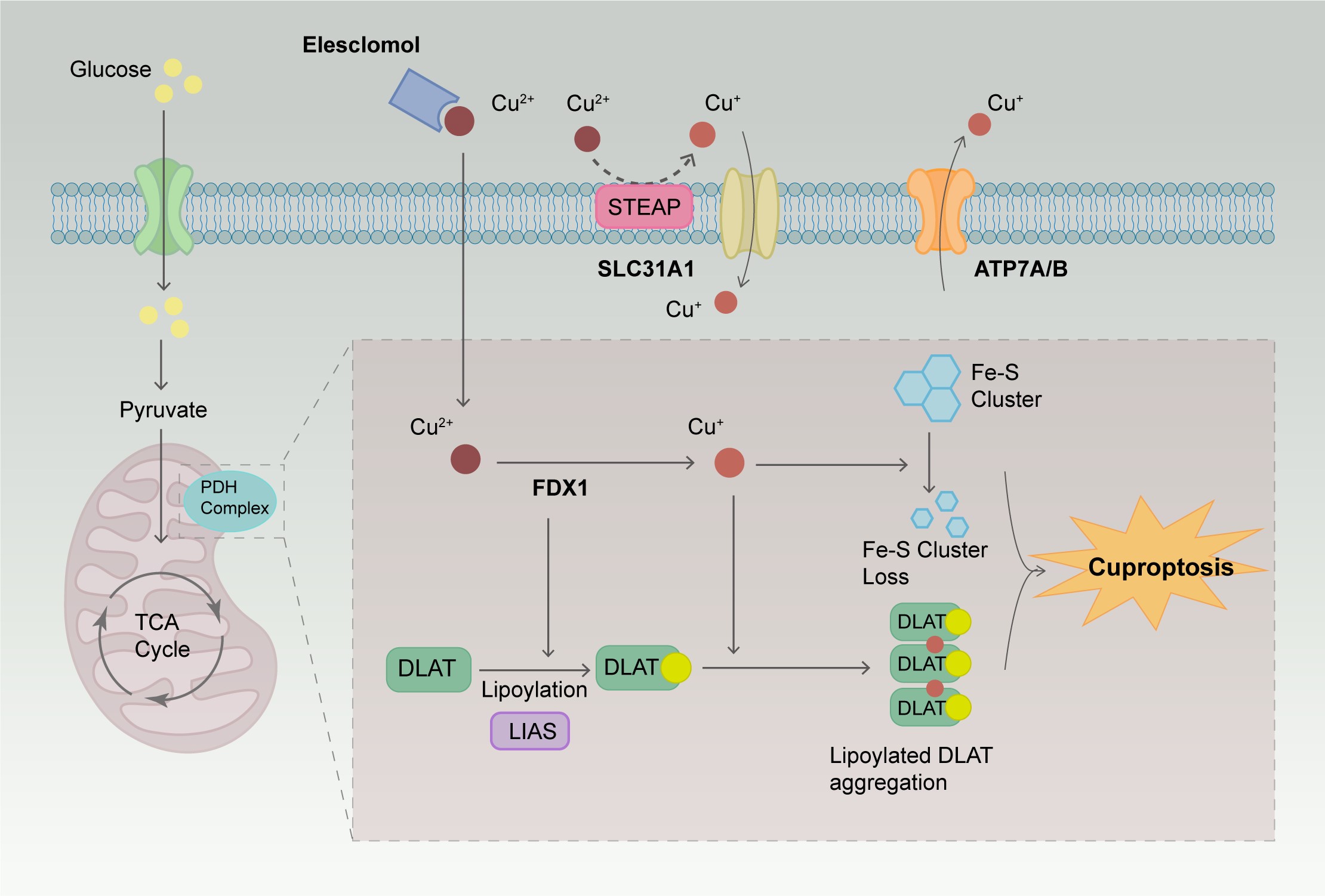 Fig. 1.
Fig. 1.
The mechanism of cuproptosis. ES can bind Cu2+ outside the
cell and transport them into the cell. When an excess amount of Cu2+ enters
the cells, it is transported to the mitochondria and reduced to Cu+. This
Cu+ interferes with the TCA cycle and the electron transport chain, leading
to the oligomerization of lipoylated proteins and the loss of Fe-S cluster
proteins, ultimately resulting in cell death. ES, elesclomol; TCA, tricarboxylic
acid; FDX1, ferredoxin 1; DLAT, dihydrolipoamide S-acetyltransferase; LIAS,
lipoyl synthase; STEAP, six-transmembrane epithelial antigen of the prostate;
SLC31A1, solute carrier family 31 member 1; ATP7A, ATPase
In addition to copper ionophores, copper transporters also regulate cuproptosis by influencing the intracellular influx and efflux of copper ions. Solute carrier family 31 member 1 (SLC31A1), also referred to as CTR1, directly transports Cu+ into cells, thereby increasing intracellular copper ion concentration, inducing mitochondrial dysfunction, and promoting cell death. Conversely, ATP7A primarily exports Cu+ from cells, thereby inhibiting copper-dependent cell death [59]. Various substances can also modulate the copper-dependent cell death process. For instance, mitochondrial glutathione (GSH) serves as a molecular partner for intracellular copper, slowing cuproptosis by inhibiting the lipoylation of mitochondrial respiratory enzymes and promoting DLAT oligomerization. Inhibitors of the mitochondrial succinate carrier, such as UK5099, and of electron transport chain complexes I/III, including rotenone and antimycin A, can inhibit cuproptosis by attenuating the activity of ES [60, 61].
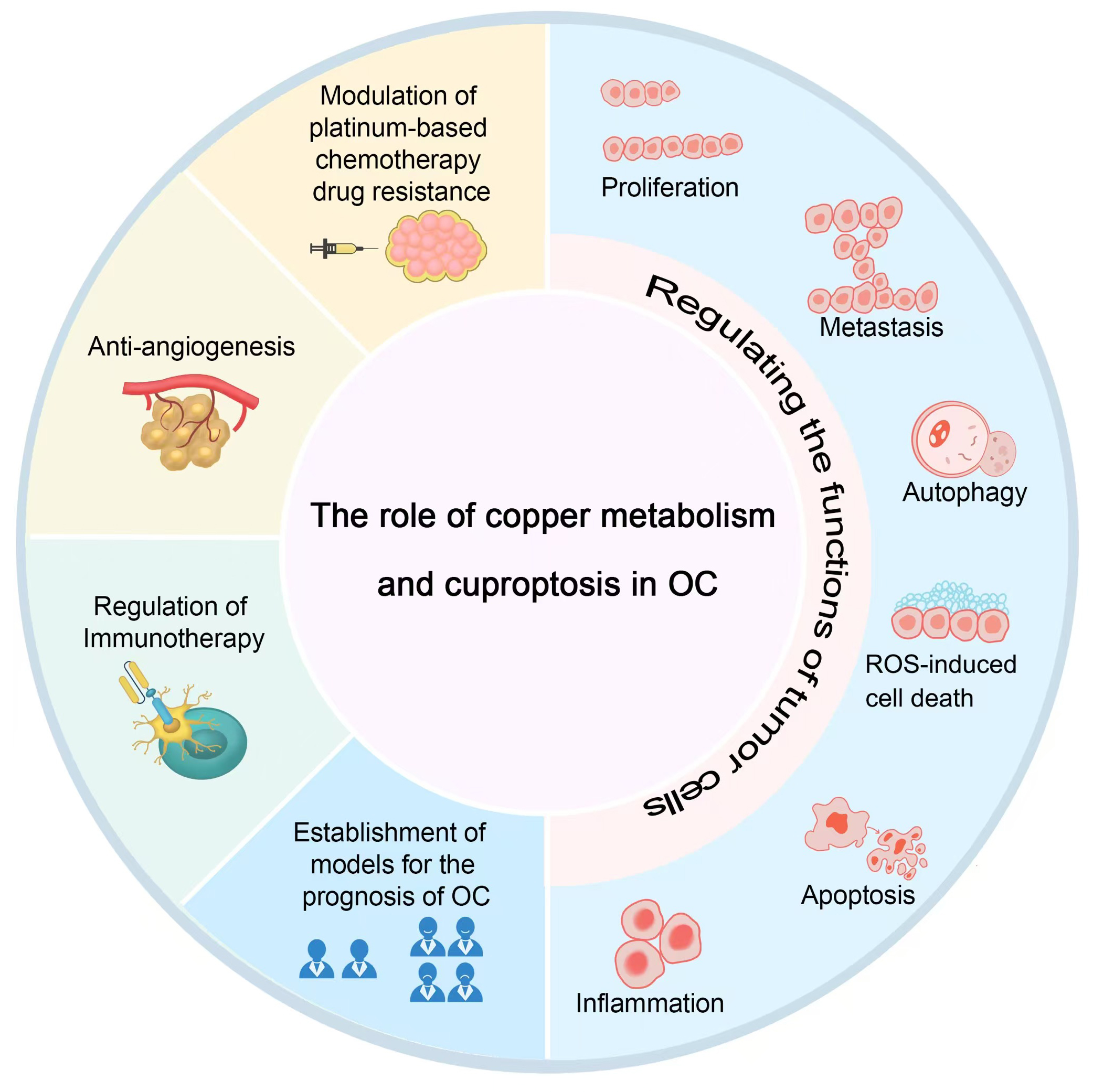
Research has demonstrated a significantly elevated copper ion concentration in OC cells compared to normal ovarian epithelial cells [6]. Cuproptosis-regulating enzymes and copper transporters play a crucial role in the onset, progression, and prognosis of OC, highlighting the therapeutic potential of inducing cuproptosis in OC cells to enhance treatment outcomes [7]. The mechanism of copper metabolism and cuproptosis in OC is described below (Fig. 2; Fig. 3).
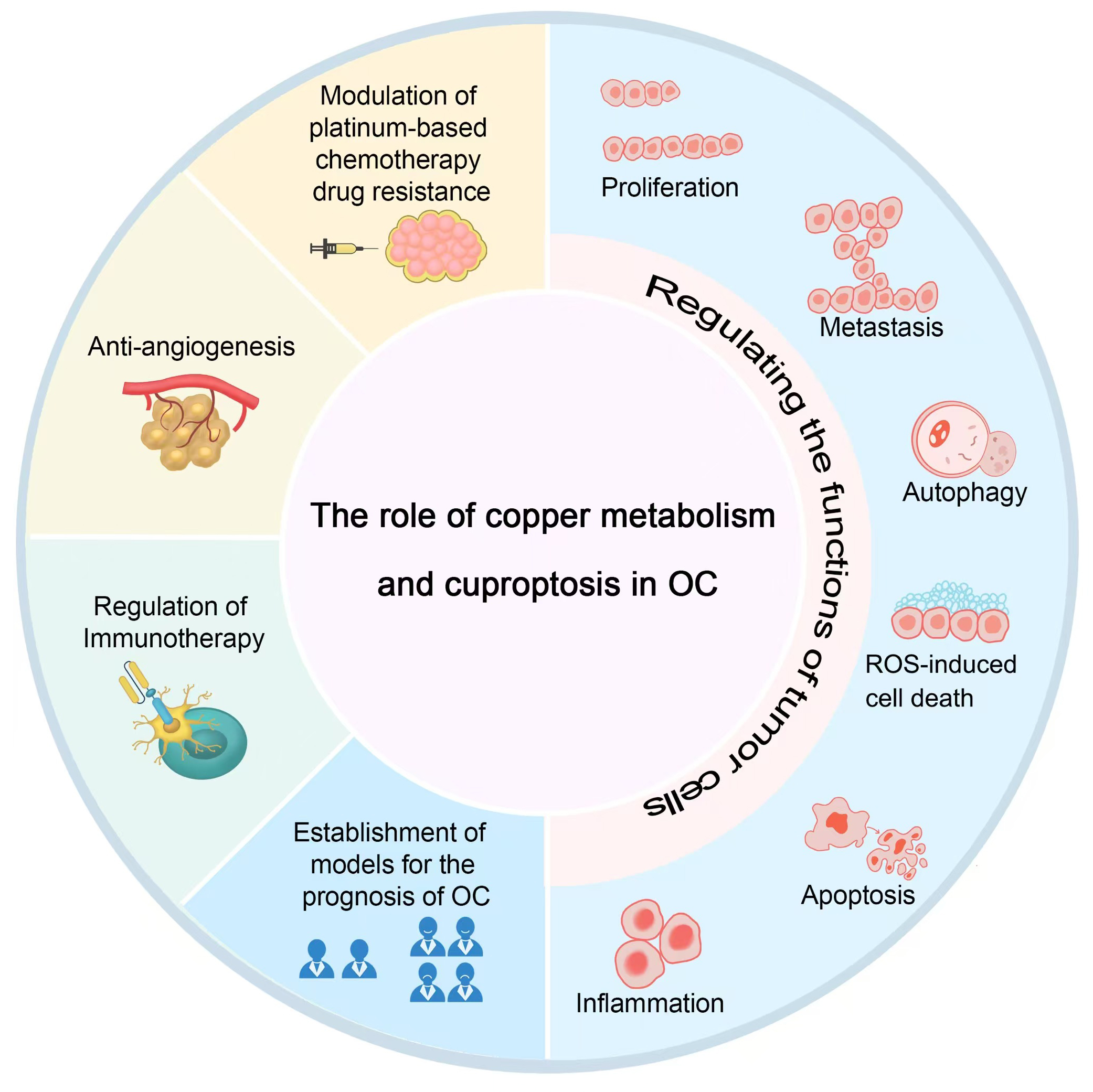 Fig. 2.
Fig. 2.
The role of copper metabolism and cuproptosis in OC. OC, ovarian cancer; ROS, reactive oxygen species. Fig. 2 was drawn using Adobe Photoshop software 2020 (San Jose, CA, USA).
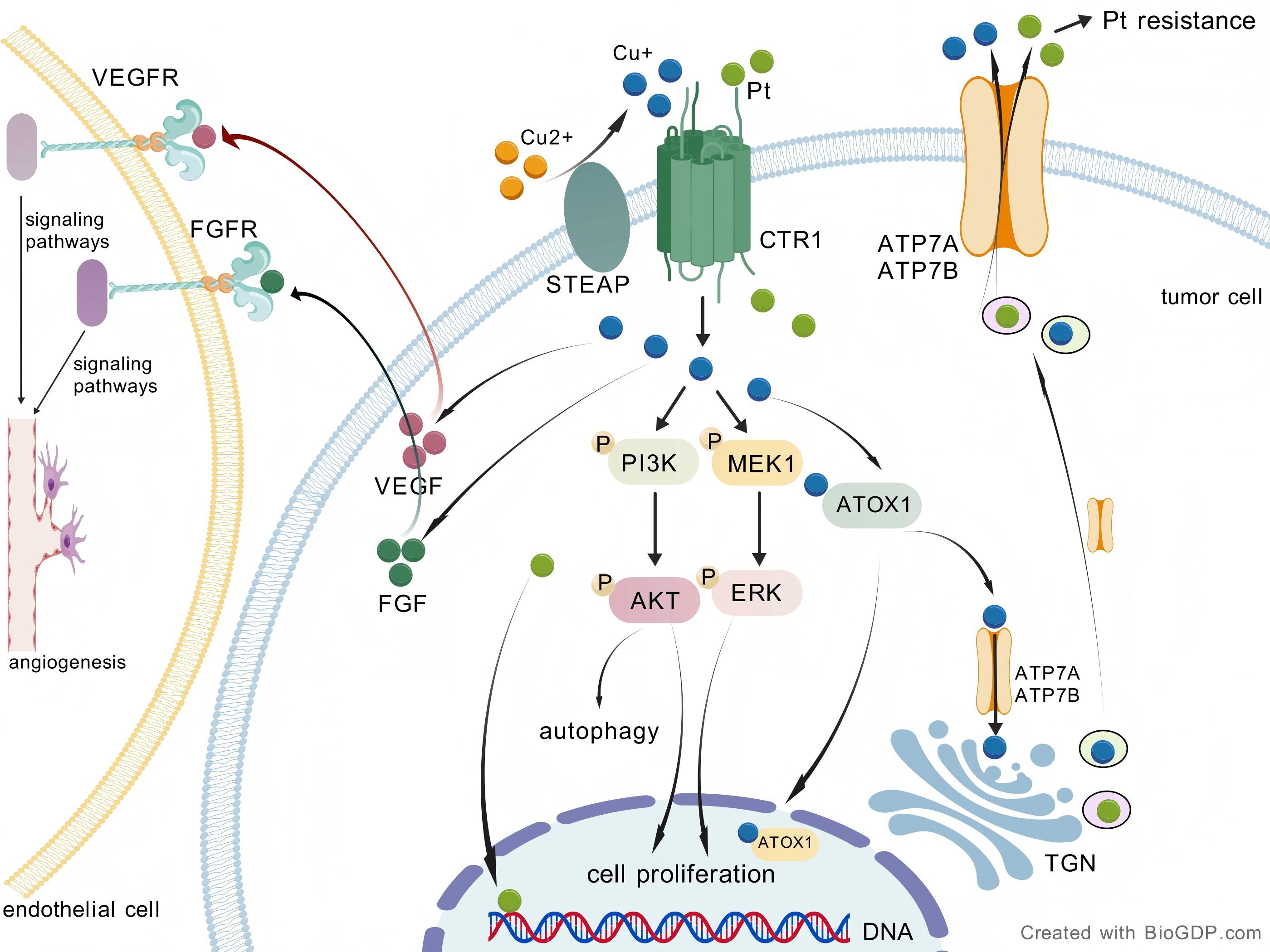
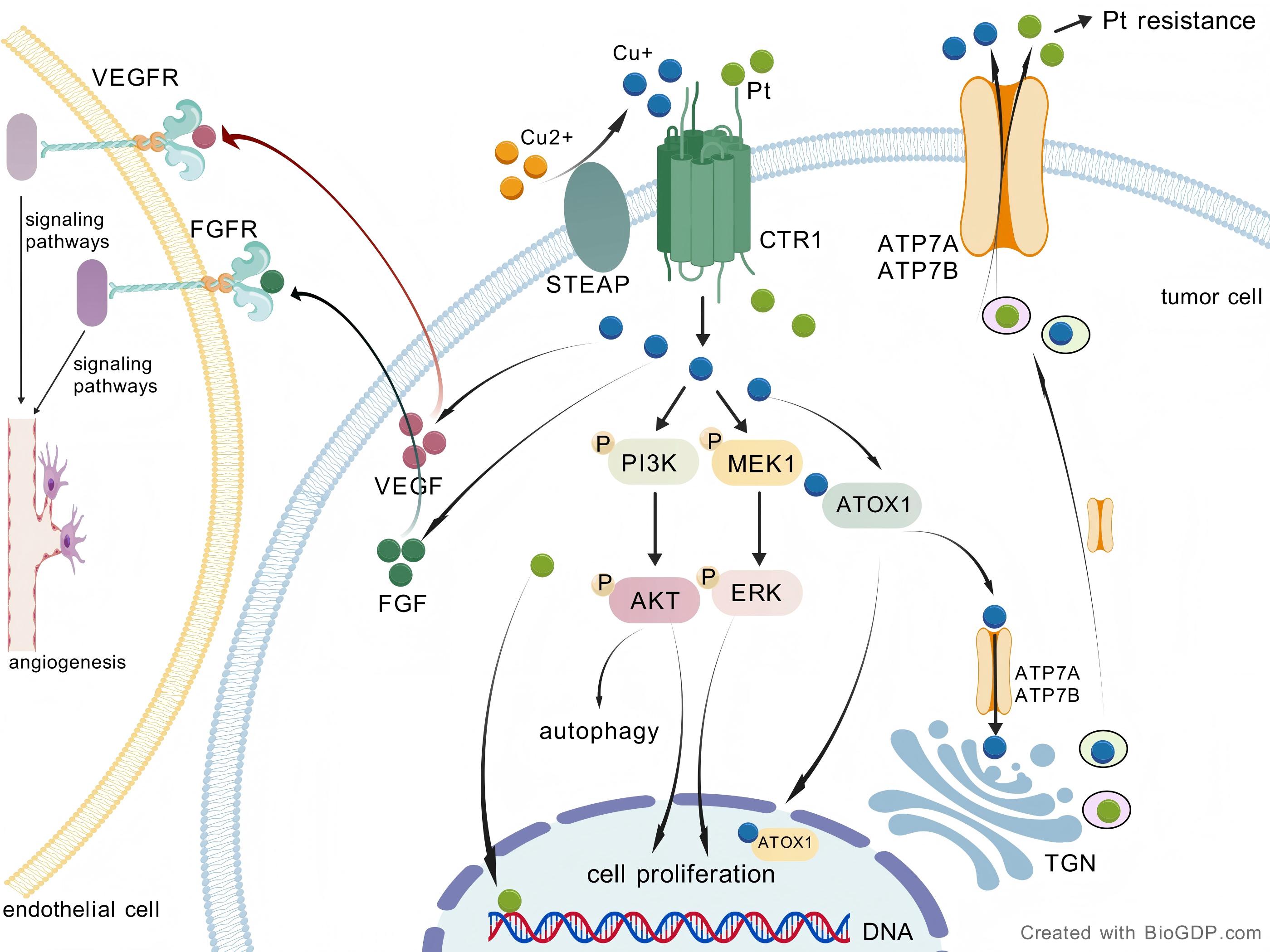 Fig. 3.
Fig. 3.
Copper-driven pathways in OC. STEAP catalyzes the reduction of
Cu2+ to Cu+, while CTR1 facilitates the transport of Cu+ into the cell. The
uptake of copper via CTR1 activates the PI3K-AKT and Ras/MAPK signaling pathways,
resulting in enhanced cell proliferation. Elevated intracellular copper levels
can modulate autophagy via the PI3K/AKT signaling pathway. Efficient efflux of
copper and platinum-based drugs through ATP7A/B transporters contributes to the
development of resistance to platinum drugs. Copper regulates the synthesis and
secretion of pro-angiogenic factors, including VEGF and FGF. After secretion from
tumor cells, VEGF and FGF bind to receptors on endothelial cells, thereby
promoting angiogenesis through downstream signaling pathways. OC, ovarian cancer;
STEAP, six-transmembrane epithelial antigen of the prostate; TGN, trans-Golgi
network; CTR1, copper transporter 1; PI3K-AKT, phosphatidylinositol
3-kinase/protein kinase B; MAPK, mitogen-activated protein kinase; MEK, MAPK/ERK
kinase; ERK, extracellular signal-regulated kinase; VEGF, vascular endothelial
growth factor; FGF, fibroblast growth factor; VEGFR, vascular endothelial growth
factor receptor; FGFR, fibroblast growth factor receptor; Pt, platinum; ATP7A,
ATPase
Elevated levels of copper ions in cancer cells can promote tumor proliferation and metastasis [63]. Cuproplasia is defined as copper-dependent cell growth and proliferation, which can contribute to tumorigenesis and excessive cellular proliferation. This process is associated with mitochondrial respiration, autophagy, redox signaling, and antioxidant defense mechanisms [64]. Copper promotes cell proliferation by activating various signaling pathways, including phosphatidylinositol 3-kinase /protein kinase B (PI3K-AKT), which may contribute to tumor development [65, 66]. Furthermore, copper uptake through CTR1 also stimulates the Ras/mitogen-activated protein kinases (MAPK) signaling cascade [67]. Knockdown or inhibition of copper chaperones such as CCS and ATOX1 significantly reduces the proliferation of cancer cells without affecting normal cell proliferation, thereby suppressing tumor progression in mouse models [68]. Autophagy, a cellular degradation process triggered by external stimuli such as starvation and hypoxia, can facilitate tumor progression. A recent study has shown that elevated intracellular copper levels can regulate autophagy, as well as the growth and survival of cancer cells, by modulating the phosphorylation of mammalian target of rapamycin (mTOR) via the PI3K/AKT signaling pathway [69].
Copper-induced cell death may occur through several mechanisms: induction of apoptosis, generation of reactive oxygen species (ROS) exacerbating oxidative damage, and caspase-independent programmed cell death [61, 70]. However, the specific mechanisms remain unclear. DSF induces apoptosis in OC cells by modulating apoptosis-related markers, including B-cell lymphoma/leukemia 2 (Bcl-2), Bcl-2-associated X protein (Bax), and caspase-3 [71]. DSF combined with copper decreases FDX1 expression, disrupts Fe-S cluster proteins, and significantly reduces OC cell viability. The combination of DSF and copper induces the production of ROS, which in turn activates c-Jun N-terminal kinase (JNK), and promotes cell apoptosis [72]. Research has also shown that copper is involved in epithelial-mesenchymal transition (EMT), an early step in metastasis that endows cancer cells with the ability to migrate and invade [73, 74]. DQ-Lipo/Cu induces cell death in ARID1A-mutant OC cells through ROS-mediated imbalance of redox homeostasis, inhibiting epithelial-mesenchymal transition, and inducing immunogenic cell death (ICD) [75]. An interaction exists between copper levels and inflammation, with serum copper acting both as a consequence and a driving factor in the inflammatory response [76, 77]. A novel copper (II)-based complex, Cu-3, can significantly inhibit the in vitro growth of SKOV3 cells and disrupt the balance of intracellular ROS levels by downregulating the expression of certain pro-inflammatory cytokines [78].
While most OC patients initially respond to chemotherapy, resistance often develops after multiple relapses, resulting in treatment failure and increased mortality rates [79]. Numerous studies have demonstrated that copper efflux transporters ATP7A and ATP7B are involved in the efflux of platinum drugs, and the expression of ATP7A/7B can affect the sensitivity of OC cells to these drugs [7, 80]. Elevated copper levels may induce the nuclear translocation of transcription factor EB (TFEB) [81], upregulating the expression of copper transporter ATP7B, which leads to platinum resistance [82]. The efficient efflux of copper and platinum drugs through ATP7A/B transporters is a crucial factor in promoting platinum-drug resistance in cancer cells [83]. Compared to platinum-sensitive cells, platinum-resistant OC cells exhibit reduced platinum accumulation, increased copper concentration, decreased total CTR1 levels, and a high correlation between total CTR1 and platinum [84]. CTR2 promotes platinum resistance in OC patients, and the CTR2/CTR1 ratio serves as an indicator for predicting platinum sensitivity in these patients. Cases exhibiting positive CTR2 expression or a positive CTR2/CTR1 ratio are associated with a poorer prognosis in OC [85]. A phase I pilot study conducted at MD Anderson Cancer Center evaluated the treatment outcomes of five patients with platinum-resistant high-grade serous OC (HGSOC) using a copper-lowering agent and carboplatin. The results provide preliminary clinical evidence suggesting that lowering copper levels may play a role in reversing platinum resistance, warranting further clinical research [86].
In OC treatment research, inhibiting tumor angiogenesis has been proven to be an effective therapeutic strategy [87]. Tumor angiogenesis is a critical step in tumor growth and metastasis. Vascular endothelial growth factor (VEGF) is a pivotal factor in angiogenesis, significantly contributing to the production of ascites. The copper levels in OC patients are significantly elevated, increasing ascites by inducing angiogenesis. Moreover, the copper content in ascites is positively correlated with VEGF levels, as well as the clinical staging [88]. Copper ions are essential for the migration, proliferation, and differentiation of endothelial cells, all of which are critical processes in angiogenesis [89, 90]. Copper is an essential cofactor in the vascular signaling cascade. It regulates the synthesis and secretion of pro-angiogenic factors, such as VEGF and fibroblast growth factor (FGF), and directly binds to angiopoietin, modulating its affinity for endothelial cells [91]. Important copper transporters, including the CTR1 importer, ATP7A efflux pump, and metal chaperones, regulate endothelial cell migration and vascular superoxide production, thereby activating angiogenesis within narrow concentration ranges [91]. Inhibiting copper-dependent angiogenesis may be a promising strategy for combating OC and warrants further investigation.
In recent years, the rapid development of immunotherapy has introduced new
prospects for the treatment of OC. Notable examples include immune checkpoint
inhibitors (ICIs), such as inhibitors of programmed death receptor-1 (PD-1) and
its ligand programmed death-ligand 1 (PD-L1) [92]. These therapies counteract the
signals from the immunosuppressive tumor microenvironment (TME) [93]. The
interaction between PD-L1 expressed on tumor cells and PD-1 receptors on T cells
suppresses the cytotoxic activity of T cells, facilitating immune evasion by the
tumors [94]. Cuproptosis has been shown to
activate immune responses and mediate immune resistance [95]. In neuroblastoma
cells, increased expression of CTR1 leads to a higher Cu2+ content, which
activates the epidermal growth factor receptor signaling pathway, resulting in
elevated levels of PD-L1 mRNA and protein expression [94]. Cuproptosis-related
genes (CRGs) in OC have been reported to potentially influence TME, clinical
pathological features, chemotherapy response, and prognosis [96]. As illustrated
in Table 1, dihydrolipoamide dehydrogenase (DLD) is a crucial component of the
TCA cycle. The expression of DLD is linked to clinical prognosis and immune
infiltration across 33 tumor types, including OC [35]. In patients with OC, the
expression of the CRG FDX1 mRNA is usually elevated [97]. Gene Set Enrichment
Analysis (GSEA) indicates that FDX1 is significantly associated with Notch
signaling, PD1 signal transduction, and mitochondrial pathways [97]. Cuproptosis
leads to the upregulation of PD-L1 on tumor cells, which complements
Research on CRGs in OC provides guidance for patient treatment and prognosis. Zhang et al. [96] developed an OC risk prediction model based on 13 CRGs. This risk scoring model, based on expression differences in OC tissues, predicts chemotherapy effectiveness and guides drug selection. A study utilizing data from The Cancer Genome Atlas (TCGA) and the Gene Expression Omnibus (GEO) datasets identified CRGs and developed the cuproptosis-related scoring (CuRS) model, which demonstrated significant predictive value for OC prognosis [99]. OC patients were divided into two clusters (C1 and C2) based on 15 CRGs related to prognosis. C1 exhibited better survival outcomes, less advanced stages, enhanced immune infiltration, sensitivity to immunotherapy, and enrichment in TCA-related pathways [100]. A copper metabolism-related gene prognostic signature (CMRGPS) of 11 genes was developed to predict overall survival in OC patients, demonstrating reliable predictive ability. Based on the median copper metabolism risk score, patients were categorized into low-risk (LR) and high-risk (HR) groups, with higher survival rates in the LR group, correlated with tumor immune-related pathways and better response to immunotherapy [101]. In some studies, long non-coding RNAs (lncRNAs) regulating cuproptosis processes were used as prognostic prediction models, offering new insights into OC prognosis [102, 103, 104]. Four cuproptosis-related lncRNAs (CRLs)—AP004609.3, AP003392.3, AP001372.2, and AC021851.1 were utilized to construct an OC risk model. Based on the median risk score value, patients were classified into LR and HR groups, with the LR group demonstrating significantly higher overall survival rates [102]. An analysis of the TCGA-ovarian serous cystadenocarcinoma (OV) dataset included 5 cuproptosis-related lncRNAs in a Cox proportional hazard model. Kaplan-Meier curve analysis revealed a poorer prognosis in the HR group. Immune infiltration analysis revealed a negative correlation between HR scores and the presence of immune-related cells [103]. Data on OC gene expression and clinical features were obtained from the TCGA, International Cancer Genome Consortium (ICGC), and GEO databases. They were categorized into 4 groups based on copper death-related lncRNAs, each exhibiting distinct survival times, immune characteristics, and somatic mutations. The 10 lncRNAs linked to prognostic assessment exhibited significant correlations with OC prognosis, indicators of the immune microenvironment, and sensitivity to the chemotherapy drug paclitaxel [104].
Recognizing the significance of copper metabolism and cuproptosis in OC, the strategy of targeting these processes for treatment has garnered substantial interest among researchers. In this context, we reviewed relevant studies on treatment strategies for OC, including copper ionophores, copper chelators, and nanotechnology, as summarized in (Table 2, Ref. [39, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146]). These approaches, as promising methods for managing OC, are gaining increasing attention.
| Drugs | Mechanism | References | Trial ID | Status | |
| Copper ionophores | |||||
| ES | Enhances oxidative stress, leading to the death of OC cells | [105, 106, 107, 108] | NCT00888615 | Completed | |
| DSF | Induces cancer cell death and enhances sensitivity to chemotherapy by promoting oxidative stress. Furthermore, it improves resistance to PARP inhibitors by modulating the DNA damage repair pathway | [107, 109, 110, 111, 112, 113] | / | / | |
| Clioquinol | Inhibits tumor growth and induces apoptosis | [114, 115, 116, 117] | / | / | |
| Copper Chelators | |||||
| Tetrathiomolybdate | Inhibits tumor proliferation and angiogenesis, prevents metastasis, suppresses immune evasion, and improves chemotherapy sensitivity | [118, 119, 120, 121, 122, 123, 124, 125] | / | / | |
| D-penicillamine | Inhibits angiogenesis and tumor growth | [126, 127, 128] | / | / | |
| Trientine | Suppresses angiogenesis and resensitizes platinum-resistant cancer cells | [129, 130, 131, 132] | NCT03480750 | Completed | |
| Cu-Based NPs | Increase oxidative stress and induce apoptosis. CuS NPs induce apoptosis through photothermal therapy | [133, 134, 135, 136, 137, 138, 139, 140] | / | / | |
| Others | |||||
| [Cu(C(13)H(14)NO(3)S)(2)](2) (Cu-A) | Suppresses cell proliferation, induces apoptosis, inhibits migration and metastasis, and inhibits angiogenesis by regulating the VEGF/VEGFR2 signaling pathway. Decreases the expression of Bcl-2 while upregulating the expression of Caspase-9 and Bax to induce apoptosis in tumor cells | [141] | / | / | |
| CPT8 | Inhibits the migration and invasion of OC cells. It suppresses angiogenesis | [142] | / | / | |
| Theaflavin-3, 3′-digallate | Enhances chemotherapy sensitivity through regulating genes related to cuproptosis | [143] | / | / | |
| Anisomycin | Inhibits the transcriptional activation of key genes related to cuproptosis in OC stem cells | [39, 144] | / | / | |
| CinOD-Cu | Effectively generates abundant hydroxyl radicals (·OH), inducing oxidative stress in the mitochondria, which results in OC cell death | [145] | / | / | |
| Cu(II)NP, Cu[Fe(CN) 5 NO] | Induces the self-generation of hydrogen peroxide and produces peroxynitrite (ONOO⁻) at the tumor site, which leads to OC cell death | [146] | / | / | |
Clinical trials information was obtained from the public database (http://www.clinicaltrials.gov/), accessed on 2 December 2024. OC, ovarian cancer; ES, elesclomol; DSF, disulfiram; NPs, nanoparticles; PARP, Poly (ADP-ribose) polymerase; VEGF, vascular endothelial growth factor; VEGFR, vascular endothelial growth factor receptor; Bcl-2, B-cell lymphoma/leukemia 2; Bax, Bcl-2-associated X protein.
ES is an injectable small molecule compound that can be used either alone or in combination with other anticancer drugs. ES binds to Cu2+ in serum, which is then effectively absorbed by cancer cells. Inside the cells, the copper in the complex undergoes redox reactions, reducing Cu2+ to Cu+, thereby generating ROS and inducing oxidative stress in the mitochondria [105]. Cancer stem cells (CSCs) that are highly dependent on mitochondrial metabolism, such as those found in OC, exhibit higher sensitivity to ES [106]. ES reduces the spheroid formation of OC stem cells (OCSCs) and lowers the expression of their markers, such as CD133 and aldehyde dehydrogenase (ALDH) [107, 108].
Compared to cisplatin alone, the combination of ES with cisplatin enhances ROS production and oxidative stress, promoting OC cell death [107]. However, a phase II multicenter clinical trial investigating the use of ES in combination with paclitaxel for refractory OC revealed that the anticancer efficacy was suboptimal, despite demonstrating good tolerability [147]. Elevated levels of lactate dehydrogenase (LDH) in hypoxic conditions are linked to more aggressive tumors. ES is more effective in non-hypoxic conditions as it disrupts the metabolic processes of oxygen-dependent tumor cells [148].
DSF has shown significant anti-cancer potential by targeting various human malignancies [149]. A study indicates that tumor cells with high ALDH activity, or ALDH-positive cells isolated from primary tumors, possess enhanced tumorigenicity and migratory capabilities. Elevated ALDH levels in certain CSC populations contribute to resistance against conventional chemotherapy [109]. DSF can sensitize ALDH+ ovarian platinum-resistant stem-like cells to platinum treatment by inhibiting ALDH activity and inducing cell death [110]. In mouse models of OC xenografts, DSF combined with copper gluconate significantly reduced tumor volume and improved survival rates [71]. Compared to carboplatin monotherapy, the combination of DSF and carboplatin enhances OC cell death by elevating ROS and oxidative stress [107]. DSF has also demonstrated efficacy in in vivo models of recurrent OC post-surgery and chemotherapy [107]. Mothers against decapentaplegic homolog 3 (SMAD3) promotes OC progression by enhancing cell proliferation, migration, invasion, and inhibiting apoptosis [110]. When combined with cisplatin, DSF synergistically inhibits OC growth by downregulating SMAD3 [111]. PARPis have demonstrated significant benefits in the maintenance therapy of OC patients, particularly those with BRCA mutations. However, resistance to PARPis may also develop in some OC patients [112]. The combination of PARPis with DSF significantly increases the expression of the DNA damage marker gamma-H2AX (gH2AX) and induces more PARP cleavage. Additionally, DSF inhibits the expression of genes involved in DNA damage repair pathways. These findings suggest that DSF can enhance the sensitivity of OC patients to PARPis through modulation of the DNA damage repair pathway [113]. Although the multifaceted actions of DSF highlight its significant potential in treating OC, clinical research involving solid tumors has not yet yielded successful outcomes [150].
Clioquinol
was initially synthesized as an antibacterial agent for the treatment of
shigellosis and intestinal amoebiasis between the 1950s and 1970s [151]. It was
later investigated in various diseases, from Alzheimer’s disease to cancer [152].
Initial study evaluating clioquinol as an anti-tumor drug showed that it
reduced survival rates by inducing apoptosis in eight different cancer cell
lines, and inhibiting tumor growth of mouse OC xenografts [114]. Another study
found that the synergistic effect of clioquinol and docosahexaenoic acid (DHA)
could inhibit the activity of the OC cell line A2780, mediated by Peroxisome
Proliferator-Activated Receptor
Tetrathiomolybdate (TM) is a rapidly absorbed and safe copper chelator with high specificity for copper binding [118]. TM exerts anticancer effects by inhibiting angiogenesis, suppressing tumor proliferation, preventing metastasis, and inhibiting immune evasion of cancer cells [119, 120, 121, 122]. Furthermore, it enhances the effectiveness of chemotherapy drugs such as cisplatin [123, 124]. TM enhances the cytotoxicity induced by doxorubicin and modulates key regulators of apoptosis in OC cells, including PARP, caspases, JNK, and p38 MAPK, which are associated with increased ROS production. TM also sensitizes OC cells to cytotoxicity induced by paclitaxel, gefitinib, and 5-fluorouracil [125]. Although TM may cause side effects such as anemia and neutropenia, these are usually reversible [154, 155, 156].
D-penicillamine, a metabolic degradation product of penicillin antibiotics, possesses strong copper-binding capabilities and can also chelate other divalent cations, such as nickel, zinc, and lead [74]. Studies have demonstrated that D-penicillamine inhibits angiogenesis and human endothelial cell proliferation by chelating copper, thereby affecting blood vessel formation and reducing tumor growth [126, 127]. Additionally, D-penicillamine upregulates specificity protein 1 (Sp1) to promote the expression of the human copper transporter 1 (hCtr1). The overexpression of Sp1 induces the translocation of p53 from the nucleus to the cytoplasm, resulting in its degradation through ubiquitination and the subsequent inhibition of ATP7A. This simultaneous regulation of ATP7A and hCtr1 by D-penicillamine can enhance the therapeutic efficacy of platinum-based drugs in cisplatin-resistant cervical cancer, particularly in phenotypes characterized by downregulation of hCtr1 and overexpression of ATP7A [128]. While D-penicillamine is commonly used to treat Wilson’s disease, it is often associated with severe adverse events (AEs) [157]. Currently, there is limited research data on the application of D-penicillamine in OC.
Trientine demonstrates superior tolerability compared to D-penicillamine and is considered a first-line treatment for Wilson’s disease [158]. Current research indicates that trientine may also have potential as an anti-cancer drug. Its chelating action inhibits copper’s role as a co-factor in angiogenesis, leading to reduced interleukin-8 (IL-8) production and suppression of angiogenesis in hepatocellular carcinoma cells [129]. A study using mouse xenograft models have identified an interaction between trientine and X-ray irradiation that inhibits fibrosarcoma growth [130]. The p38 MAPK pathway appears to play a significant role in trientine-induced apoptosis in fibrosarcoma cells [131]. Copper-lowering agents can resensitize platinum-resistant cancer cells by enhancing platinum uptake mediated by CTR1. A dose-escalation study of trientine combined with carboplatin and pegylated liposomal doxorubicin in first-recurrent epithelial ovarian, fallopian tube, and peritoneal cancer demonstrated good tolerability and high safety. However, it showed limited anti-tumor activity, with no correlation observed between clinical response and serum copper levels [132].
Nanotechnology provides an innovative strategy for early diagnosis, prevention, and personalized treatment of cancer through the application of NPs and quantum dots.
NPs can serve as direct anti-tumor therapeutic agents or as carriers to enhance controlled drug delivery, thereby improving efficacy and reducing side effects [159]. Copper NPs, including Cu2O, CuO, and zero-valent copper (CuNPs), have rapidly advanced in recent years and have emerged as some of the most promising candidates for treating various cancers, including OC [133, 134]. The particle size of CuNPs typically ranges from 1 to 100 nanometers and can be synthesized using various techniques, including chemical reduction, physical methods, and biological synthesis [133]. These NPs exhibit significant potential in cancer treatment due to their interactions with malignant cells. Their small size and large surface area facilitate efficient penetration of cell membranes. Upon entering the human body, CuNPs release ROS, which increase oxidative stress and ultimately induce apoptosis in cancer cells [135].
Research on CuNPs in OC has garnered increasing attention. Copper sulfide NPs (CuS NPs), due to their unique optical properties, small size, low production cost, and low cytotoxicity, are a promising new nanomaterial for cancer photothermal therapy (PTT). CuS NPs target tumor cells and enter the cell nucleus, where subsequent activation by near-infrared laser irradiation leads to increased temperature in the nucleus, resulting in tumor cell apoptosis [136]. Feasibility study using CuS NPs for OC PTT guided by positron emission tomography (PET) and magnetic resonance temperature imaging (MRTI) have demonstrated significant tumor ablation in an OC mouse model, with minimal impact on surrounding healthy tissues [137]. Folate receptors, which are overexpressed on the surface of OC cells, make them ideal candidates for specific targeting through functionalized NPs and other contrast agents. FA-CuS NPs have been shown to be an ideal contrast agent capable of accurately identifying circulating ovarian tumor cells in flow cytometry using photoacoustic flow cytometry, providing an opportunity for precise detection of early cancer metastasis [138]. The integration of nanotechnology with traditional herbal medicine, aimed at enhancing the precision of targeting specific areas and improving the efficacy of herbal treatments, has emerged as a prominent therapeutic strategy [139]. CuNPs synthesized from Camellia sinensis leaf extract show excellent anti-OC properties and hold potential as novel chemotherapy adjuvants [140]. Due to their small size and unique surface properties, CuNPs exhibit low toxicity and compatibility with biological systems [135]. However, the safety of CuNPs remains under investigation. There is a scarcity of animal studies and preclinical trials concerning CuNPs in OC, highlighting the urgent need for more high-quality research to explore and validate their potential in the future.
Copper-based complexes have emerged as promising alternatives to traditional platinum-based anticancer agents [160]. A novel Cu2+-based complex, Cu-A, inhibits the proliferation of SKOV3 cells and human umbilical vein endothelial cells (HUVECs), induces cell apoptosis, and suppresses angiogenesis by regulating the VEGF/VEGFR2 signaling pathway [141]. The low-toxicity Cu2+ complex CPT8 significantly inhibits the migration and invasion of OC cells and demonstrates notable anti-angiogenic activity, as evidenced by its effects on tube formation and sphere germination in HUVECs [142].
Currently, researchers have sought to identify natural molecules with potential applications in oncology. Theaflavin-3,3′-digallate (TF3) is a polyphenol present in black tea, which exhibits lower cytotoxicity to normal ovarian cells than to OC cells. TF3 treatment significantly reduces GSH levels and upregulates CTR1 protein levels in OC cells, enhancing their sensitivity to cisplatin [143]. Anisomycin is an antibiotic derived from gray mold that inhibits peptide bond formation and protein synthesis by binding to the 60S ribosomal subunit [144]. The study has shown that Anisomycin significantly suppresses the activity of OCSCs. Anisomycin inhibits the transcriptional activation of key genes in the fatty acid pathway, including FDX1, DLD, DLAT, and PDHB, which may potentially lead to cuproptosis in OCSCs [39].
Chemodynamic therapy (CDT) employs agents-catalyzed decomposition of
hydrogen peroxide (H2O2) into highly toxic hydroxyl radicals (OH),
thereby inducing cell death [161]. However, the efficiency of
Through a comprehensive literature review, this article systematically elucidates the intrinsic relationship between copper metabolism, copper-induced cell death, and OC. Additionally, it explores various copper-related treatment strategies in the context of OC, thereby offering novel insights for potential therapeutic approaches to this malignancy. Copper-based therapies, particularly those involving copper-based nanotechnology, represent a promising emerging strategy for OC patients.
To date, there have been limited in vivo trials investigating copper-related approaches for the treatment of OC, and the safety of these strategies remains uncertain. Furthermore, the efficacy of copper-based treatment modalities across various types of OC has yet to be clearly established. Organoid models can be developed to predict the drug sensitivity of copper-based therapeutic strategies across various subtypes of OC. Further research involving OC animal models is necessary to evaluate the safety of these copper-based treatment strategies. Ongoing scientific advancements and innovation are expected to lead to more precise and effective therapeutic approaches for OC patients, ultimately improving treatment outcomes.
QW, HD, and JS made substantial contributions to the figure drafting, design, and data visualization critical to the manuscript’s scientific presentation. QZ conceived and designed the structure of the article. YX reviewed the literature and wrote the manuscript. All authors made editorial revisions to the manuscript. All authors have read and approved the final version of the manuscript. All authors have participated sufficiently in the work and agreed to be accountable for all aspects of the work
Not applicable.
We would like to express our sincere gratitude to all those who assisted us during the preparation of this manuscript. We also extend our thanks to the peer reviewers for their valuable opinions and suggestions.
This work was supported by the Medical Science and Technology Project of Zhejiang Province (No. 2023KY1250, 2023KY362) and the Scientific research project of Shaoxing Municipal Health Commission (No. 2023SKY041).
The authors declare no conflict of interest.
We acknowledge that during the preparation of this work, the authors utilized ChatGPT-4.0 for spell and grammar checking. Following the use of this tool, the authors reviewed and edited the content as necessary and assume full responsibility for the final publication.
References
Publisher’s Note: IMR Press stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.